TARTU RIIKLIKU ÜLIKOOLI IMETISED УЧЕНЫЕ ЗАПИСКИ ТАРТУСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА ACTA ET COMMENTATIONES UNIVERSITATIS TARTUENSIS ZOOLOOGIA-ALA
|
|
|
- Marko Kirss
- 4 aastad tagasi
- Vaatused:
Väljavõte


1 TARTU RIIKLIKU ÜLIKOOLI IMETISED УЧЕНЫЕ ЗАПИСКИ ТАРТУСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА ACTA ET COMMENTATIONES UNIVERSITATIS TARTUENSIS ZOOLOOGIA-ALASE ID TOID ТРУДЫ ПО ЗООЛООГИИ


2 TARTU RIIKLIKU ÜLIKOOLI TOIMETISED УЧЕНЫЕ ЗАПИСКИ ТАРТУСКОГО ГОСУДАРСТВЕННОГО УНИВЕРСИТЕТА ACTA ET COMMENTATIONES UNIVERSITATIS TARTUENSIS ALUSTATUD 1893.a. VIHIK 408 ВЫПУСК ОСНОВАНЫ В 1893.г. ZOOLOOGIA-ALASEID TÖID ТРУДЫ ПО ЗООЛООГИИ XI Põder Eestis Лось в Эстонской ССР II TARTU
 продолжают тему биологии лося, начатую в предыдущем выпуске (В 407 ). Teil.I Tartu Riikliku ülikooli To leetleed, 1977. 4fifi. Цит.: Уч. зап. Тартуск.")
3 Bedakteloonlkolleeglue: J. Aul (raet. tolo.), H. Ling, H. Ream. KeoHolevae TRÜ Toloetietoe nr. 408 toodud ortlklitee Jatketakae TRÖ Tolcetietee nr. 407 aluetetud põdra bioloogia kieltluat. ' Статьи настоящего тома "Ученых записок ТГУ" (вып. 408 ) продолжают тему биологии лося, начатую в предыдущем выпуске (В 407 ). Teil.I Tartu Riikliku ülikooli To leetleed, fifi. Цит.: Уч. зап. Тартуск. гос. ун-та, 1977, Ref. 1 Aota et coomentat. Univere. Tartuenele, 1977, 408. Tsrlu Riiklik ülikool. I977


4 podbapopulatsiooni stbuktuub JA DUtAAMKA EESTI HSV- s Populatsiooni süsteemse analüüsi katse H, L l n g POPULATSIOONI SOTSIAALNE STRUKTUUR Sotsiaalse struktuuri algühikuke oa koos esinevate ja koos tegutsevate isendite grupp. Jagamatuteks elementideks, millest koosneb sotsiaalse struktuuri algtihik, on soolt ja vanuselt erinevad isendid. Ka sel juhul, kui grupi moodustavad sama soo ja vanusega isendid, pole tegu lihtsa elementide summaga, vaid kvaliteedilt uue ühikuga, kus avaldub grupiefekt ja mille osa populatsiooni sotsiaalses struktuuris on erinev vsrreldes sellega, kui samad isendid esineksid üksikutena. Grupi koosnemist erinevatest elementidest nimetame grupi kompositsiooniks. Grupi koostisest iffigime sel juhul, kui analüüsime gruppi moodustavate eri elementide protsentuaalset vahekorda. Tingituna materjalide kogumise meetodist oli võimalik eristada ainult kolm kvalitatiivselt erinevat rühma: vanad pullid, lehmad ja vasikad.* Järelikult vsiks grupis esineda maksimaalselt kolm erinevat elementi, elementide ühikute (isendite) arv vsiks aga olla märksa erinevam. Grupi kompositsiooni tähistamiseks kasutati kolme arvulist tähistust, kus järjekord määrab elemendi tüübi (pull, lehm, vasikas) ja arv antud tüüpi kuuluvate isendite arvu. Mingi elemendi puudumist grupis tähistab " - Näiteks grupi kompositsioon, kus esineb üks pull, kaks lehma ja kolm vasikat, edasi antud valemiga "1 2 3", lehmast ja vasikast koosnev on * Kompositsiooni kaueaalanalüüeil kasutati ka aastal Mahtras kogutud materjale, kus oli eristatud viie rullina (vt. lk. 19). 3

 koosesinemine ühest lehmast ja vasikast (vasikatest) koosnevate gruppidega (komponentidega) on tähistatud järgmiselt:» }2<? t 1gj\". Ja T a b e l?")
5 grupp aga Tulemiga 1 1". Sagedamini kooe eelnevaid elemente, mla eelnevad paijudee gruppides (näiteks lehm vasikaga), on nimetatud komponentideks. Kompositsioonide komponentide sageduste võrdlusel on siiski otstarbekam soo Ja vanuse tähistamiseks kasutada üldtunnustatud tähistusi (<?, j [juv.j ), märkides vaetava tähistuse ette isendite arvu ning eraldades võrreldavad kompositsioonid kooloniga. Hälteks kahe ja enama pulliga gruppide (komponentide) koosesinemine ühest lehmast ja vasikast (vasikatest) koosnevate gruppidega (komponentidega) on tähistatud järgmiselt:» }2<? t 1gj". Ja T a b e l? 4 põdra sotsiaalse struktuuri selgitamiseks analüüsitud is end liiste gruppide arv kuude ja aastate lõikes \>Kuu Aastefx X XI XII Kokku З ь Kokku


6 uu* Ott Ott /зеьчза at? tau 1Ш то 1171 Joon. 27. Põdrapullide esinemissagedus laendilletes gruppides oktoobrikuul aastatel Arvestades seni teadaolevaid andmeid normaaleelt koos esinevate ja koos tegutsevate põdragruppide suurusest, tuleb suuri gruppe vaadelda kui agregataioone, ais moodustuvad teatud tingimustel lühemaks ajaks ning mille УЯу mete vahel pole sotsiaalses struktuuris Iseloomulikke suhteid. Sel põhjusel kasutati analüüsil vaid isendiliete gruppide andmeid, mis sisaldavad kõigist populatsiooni isendeist oktoobris 83,1...95,1, novembris 83,2...93^8 Ja detsembris 77,9...89,4 56. Välja jäeti aasta andmed. Kokku analüüsiti 7935 grupi struktuuri (tabel 34). 5

7 I г k/y Joon. 28. Põdialehmade esinemissagedus isendilistes gruppides oktoobrikuul aastail Ф л ' Jf // у/ 7 \ \ Xx N is/gr Joon. 29. Vasikate esinemissagedus isendilistes gruppides oktoobrikuul aastail G
 on kõige muutlikum olnud 1- Ja 5-ieendilistes gruppides, kõikudes vastavalt 13,5...32 % Ja 3,6.")

8 4.1. KLSEBETIDg BSUHCSK SXGEDDS ISBTDILISTKS QHtJPPIDKS OKTOOBRIKUUL Pullide esinemissagedus eri aastatel (Joon. 27) on kõige muutlikum olnud 1- Ja 5-ieendilistes gruppides, kõikudes vastavalt 13, % Ja 3,6...18,8 * rahel, ülejäänud gruppides on pullide esinemissagedus olnud suhteliselt stabiilne ja kui jätta iga eri suurusega grupi puhul arvestamata ainult ühe aasta andmed, siis esinemissagedus kõiguks 2,5 kuni 4 % ulatuses. ainult Arvestades ka eri aastail analüüsiks kasutada olnud andmete hulka, on vaadeldaval ajavahemikul peaaegu pidevalt vähenenud nullide osa a 3-1 sendilist es ning suurenenud 4-,1a 5=1аащщц даавдаая- Ighaade esinemissagedus eri aastail (joon. 28) on kõige muutlikum olnud samuti 1- ja 5-isendilistes ning stabiilseim 3-isendilistee gruppides. Enamikul aastatel on aga lehmade osa üksikute hulgas olnud väiksem kui pullide osa ning ainult ajavahemikus (madala viljakuse periood!) oli see suhteliselt suur, 4- ja 5-isendiliste gruppide osa same ajavahemikus oli aga suhteliselt väiksem. Seega põdralahmede esinemissagedus eri suurusega rühmades sõltub antud aasta populatsiooni viljakuse tasemest: suure vll-lakuse puhul on vähem lehmi üksikuina Ja rob*» i«hnri i-,1a 5-isendillsteB gruppides. väikese viljakuse puhul aga vastupidi. 2- ja 3-i sendi - Ustes gruppides olevate lehmade esinemissageduse ja populatsiooni viljakuse vahel ilmset seost ei esine. Vasikate esinemissageduse muutlikkus eri aastail (Joon. 29) on olnud suhteliselt ühesugune kõigis gruppides. Vasikatest % on esinenud 3- ja 4-isendilistes gruppides ning 2-1sendil!etes gruppides on neid alati esinenud rohkem kui 5-isendilistes, viimastes eriti vähe madala viljakuse perioodil aastal täheldatav tunduvalt väiksem esinemissagedus 3-isendilietes gruppides korrespondeerub lehmade vaikse esinemissagedusega. 7

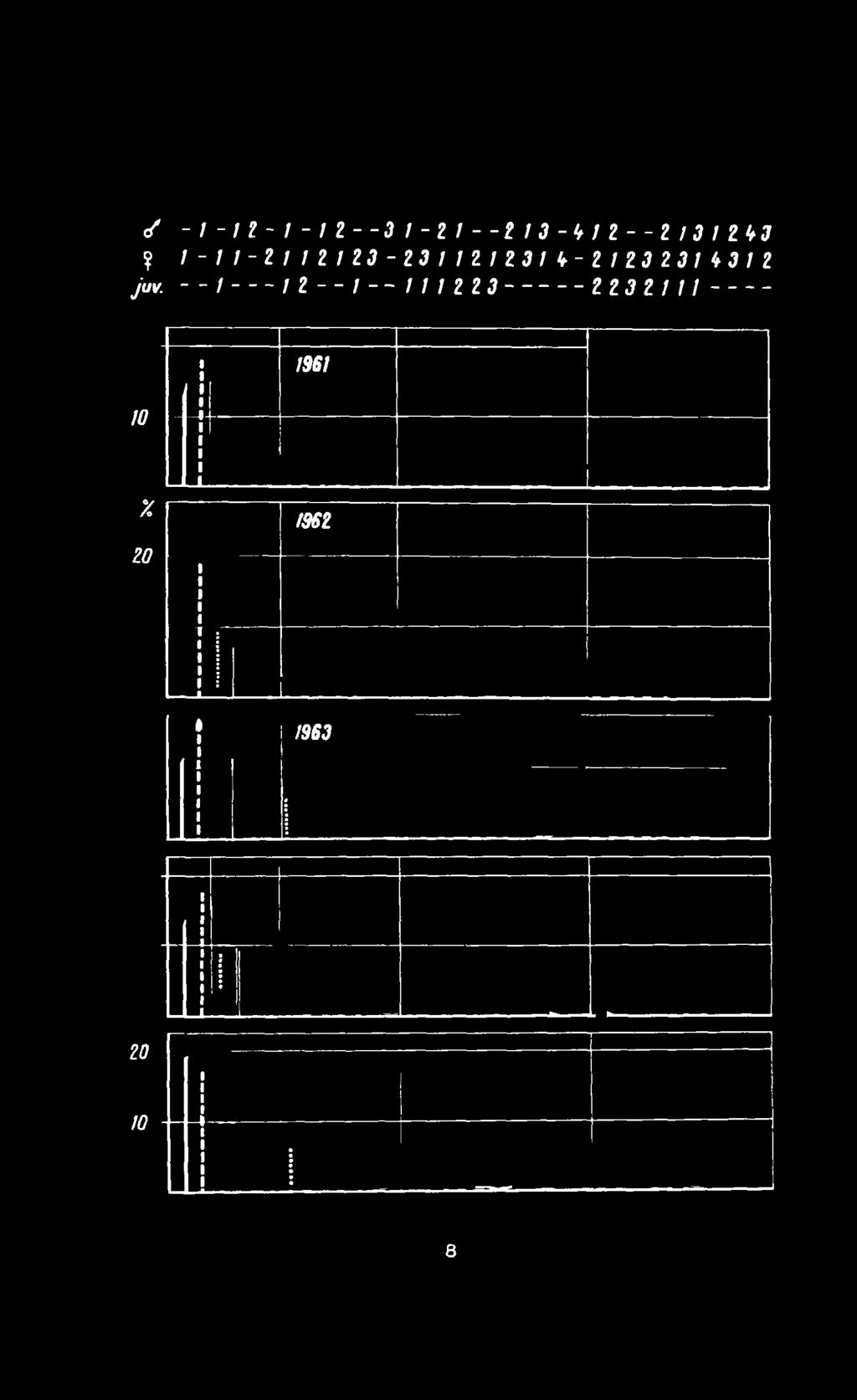
9 -/-/ -/-/2--Д /-2/--*/J-W 2--2/J/ZW? /-Z/-2//2/2J-23/Z2/23/t-2/ZJ2JZ43Z2 ju* - - I / 2 / / ZZ22J ггзгпi /56/ to X 1962 го i /963 i 25 /0 8

 oktoobris aastail 1961.")
10 ? JUV. -l-fz-l-iz-~31-zi--zi3-itl2--zi3lii,3 /-/г-гпгиз-гзг/г/гз/ь-г/гзгз/аэгг -- / / / / г гз 2 23 г 111 / st и 11 1 \ \ i * 1368 v 1 /Л 1 1 viwnv tvftv> % Ml - yf \ : / v/l VlTTv-^y tvfk К! X УГ л 1370 \ P 1 I -L- V/ IIVTK ^ T T T K ^ ^ - /,971 У/ Ут1^-^\/ТК-. Joon ,5-isendiliste gruppide erinevate kompositsioonide esinemissagedus (%) oktoobris aastail : 1 - ainult leiuaaa, 2 - ainult pulliu, 3 - pullid ja lehmad ilma vasikata, 4 - lehm ühe vasikaga, 5 - lehm 2 vasikaga. 2: 9
 (Ql>P>oj) tai >p>o.o5) (P <0.05)!» (P< 0.01) i» «(P<0.001) Joon. 32.")
11 /36/ P-% 70>p>'i-Q tto>p>zo 10 >p > 5 joon. 31. Aastate sarnasus-erinevus kompositsioonide esinemissageduste alusel (kasutatud X; menetlust). Peen joon näitab sarnasuse, jäme joon erinevuse usaldatavust. 1 + < ) (Ql>P>oj) tai >p>o.o5) (P <0.05)!» (P< 0.01) i» «(P<0.001) Joon. 32. Põdrapopula ts iooni sotsiaalse struktuuri komponentide vaheline korrelatsioon isendilistes gruppides oktoobrikuul. 10
.")

12 4.2. xm JUSSiSE SABIASUS ОИООВКШГО ISHDILISIE 6H0PPIDE KOEPOSITSIOOZIBK ESHHCSSAEEDOSE ALUSEL Andmed ksigl eri пета koapoeitsiooniga gruppide esinemleeagedueeet oktoobris aastail on eeitatud JOOB. 30. Eri aastate erinevate kompositsioonide esinemissageduste absoluutarvude omavaheline v8rdlus abil v8imaldas selgitada omavahel sarnased Ja erinevad aastad (Joon. 31). Kui naaberaastad kompositsioonide sageduselt omavahel oluliselt ei erinenud, vaadeldi teiste aastatega võrdlemisel nende andmeid koos. Kompositsioonide esinemls-' sageduselt sarnasemad aastad on , , ja , naaberaastaist ei erinenud aastad Ja , usaldatavale tasemele ei küflndinudaaetate 196З erinevused. KSige Järsemad muutused vörreldes eelneva aastaga esinesid aastail 1968 ja Seega 10 aasta Jooksul esines nii kompositsioonide esinemissageduselt sarnaseid kui ka oluliselt erinevaid aastaid, enam-vähem sujuvaid üleminekuid Ja ka järske muutusi EBI KOMPOSITSIOOIID8 ESÖEUSE SEADUSPÄRASUSED PSdragruppide tuumikuks peetakse nn. emaperekonda (mother-family) - p8dralehma tema vasika v8i vasikatega/19, 34, 91, 211/. Omaette erinevaike elementideks, kuld kes v8lvad liituda nii omavahel kui ka vasikaga emaloomaga, on pullid ja vasikata lehmad. Populatsiooni sotsiaalse struktuuri sisemiste seaduspärasuste selgitamiseks näib sobivaim olevat analüüsida just nende kolme rühma omavahelisi suhteid: erineva arvu pullide liitumist vasikaga Ja vasikata lehmaga, vasikata lehmade liitumist pullide ja vasikata lehmaga, üldiste seaduspärasuste ja eri aastail eelneda v8ivate erinevuste selgitamiseks tehti vaetav analüüs Iga aasta v8l kompoaiteioonide suhteliselt sageduselt mitteerlnevate naaberaastate summaarsete andmete alusel kae oktoobrikuu v8i aiatee aastast ka novembri Ja detsembri kohta. 11 ainult


13 T a b e l 3 5 Vasikata põdralehmade koosesinemine pullidega Aasta 3 df X 0,59 2 >0,7 196*4-1965»»,76 2 >0,05 >2j :1 cf; : >2:/; 1967 " 2,55 2 >0,3 Teoreetilisest sagedusest tunduvalt suuremad väiksemad V *o Ö 1968 " 8,74 2 >2g«0<fj 1^:0if; : 1 cf; 1969 " 2,52 2 >0,2 >2y«1<fi >2f:0cf; 1970 " 6,06 2 >2yi1<f; >2^:0(f; 1971 " 2,76 2 >0,2 >2gi>2(f; >2f$0< ; 1968 IMU 0,37 2 X>, XI 2,26 2 >0, XIX 12,05 2 >2^i>2<f; 1^;>2/;>2^:0(f; 1970 X*XI 10,34 4 2^:>2/; 1^i0if; 2^:0(^;>5^i0(/; 1970 XII 8,83 2 >2^:>2cf; 1 1>2С^;>2^:0< ; 1971 XMI1 2,43 2 >0,2 >2^1 >2/, >2^:0/ T a b e l 3 6 Erineva vasikate arvuga põdralehmade koosesinemine pullidega Aseta 5 t 2 df p Teoreetilisest suuremad aageduseat tunduvalt väiksemad i 19,00 3 * yl<$:>2 lj; 0<?i2g>2j; yi<f!^>2j; 04: >2^1j; 1964*1965 " 25,12 3 >1lfl>2j1di Orf:2?>2J; 0<f: >2^1õ;>1<fi,-^>2j; 1967 " 1,17 2 X) ,54 3 >0 05 y<sl>2ftj, 0rfl2j>2J; 0(f:>2^1 j; yi<^:2^>2j; 1969 " 27,90 3 0rfl2j>2j yltfs >2 J >1iJ :10ld; >1(^i2o>2tj; 0( l>2o1j; * * >1<#:1ofcj; 1970 " 36,38 6 )2<#i>2?1j, lrf:2^>2jj 0<f: >2^1 j; 1971 *' 52,62 3 >1<:>2(1ji 0i?l2)>2j; 0(?:>2^1 j; >1cfi2^>2j; 1968 XI+XII 5,04 2 X) 05 0^:1j>2Jj yi^l^j; ' 1969 XI 14,71 3 Jlii>2^1Ji 0if!lj>2J; 0tf;>2^1 j; >1^:1g>2«j; 19b9 XII 7,14 2 yli?!>2o1 j ; Otfl y1^>2j ; 0(f:>2^1jt yi<f:>1^>2j; 1970 X*XI b4,06 6 >2<$s>29ljs lf:)2o1ji Ojl2{>2J! 0<$:>2^1j; 1970 XII 17,5* 3 (J 1 >2^1 J; 0i#m>2ji CX?:>2^1jt>1<f: 1y>2j; 1971 XI+XII 17,80 3 >1#l>2^1ji 0<Js>2^1j гг


14 Yaaifota koosesinemine pullidega Eraldi oli võimalik analüüsida erineva arvu lehmade koosesinemise eageduei erineva arvu pullidega (tabel 35). Oktoobris esinesid juhuslikust jaotumusest usaldataval tasemel ainult ja aasta andmed, detsembri andmeist oli usaldataval tasemel erinevus ja a. Oktoobris esines juhuslikust sagedamini kompositsioon 1 1 <f v8i >2g : > 26 ning harvemini kompositsioon >2$ '.Oi - e. t. kahe v8i enama vasikata lehmaga liituvad pullid sagedamini kui üksiku vasikata lehmaga. Erandiks on ainult aasta, kus pullid esinesid sagedamini eraldi lehmadest ja harvem koos lehmadega. H o v e m b r i s - d e t s e m b r i e i l m n e b v e e l gi tugevamalt pullide afiinsus liituda suurema arvu vasikata lehmadega, kuna samal ajal ühe lehma juures neid harvem: prevaleerivad kompositsioonid >2g : > 2<f esineb : Od 1 ning juhuslikust jaotumusest märksa harvem esinevad kompositsioonid 1g :>2<f ning >2g : 0<$. ja Vasikaga psdrelehwh* koosesinemine pullidega Juhuslikust jaotumusest usaldataval tasemel erinevust ei esinenud ainult kolmel aastal ning neistki kahel oli usaldatavus 90 ja 95 % vahel (tabel 36). Enamikul aastatel oli erinevus juhuslikust usaldatav ksrgemal tasemel. К 8 i g i 1 k u u d e l esineb tunduvalt sagedamini pulle gruppides, kus on kaks lehina ühe vasikaga А ваша! aial on neid harvem gruppides, kus on kaks lehma.lakaks v8l rohkem vasikat: prevaleerivad kompositsioonid )1<f: >2^1 j ja 0<S : 2ф > 2j Pullide esinemine üksi, vasikata Ta" vasikaga pcdralehmaga Enamikul aastatel esineb juhuslikust jaotumusest usaldataval tasemel vsi selle lähedane erinevus (tabel 37). K8igil kuudel ja aastatel on kaks,1a enam pulli esinenud sagedamini kooe vasikata psdralehmadega kui esinenud kas uksi vsi vasikaga p3dralehmaga: üks pull on aga 13 sagedamini
 I 1,08 2 >0,5 1964*1965 \" 13,18 2 >2Jt>1j0j 1ifi>1gj; >2(fl>1)j! Icfsyi^Oj; 1967 8,b5 2 >2#l»1^0j 1ifl>1^jl 1966 \" «.")

15 T a b e l 5 7 Pallide esinemine üksi, vasikata ja vasikaga põdralebmaga lasti 3 и X2 dl P Teoxeetllleeet euur.osd eegedusesi tunauvelt väiksemad ) I 1,08 2 >0,5 1964*1965 " 13,18 2 >2Jt>1j0j 1ifi>1gj; >2(fl>1)j! Icfsyi^Oj; ,b5 2 >2#l»1^0j 1ifl>1^jl 1966 " «.83 2 >2<fl>1fOJ 1969 " 4,41 2 >0,1 >2^i>1^0j 1970 " 4,37 2 >0,1 >2#iyl^0j, >2/10^ " 19,37 2 >2#«>1g0j l'»>1^jl >2<fl 1)3 i 1968 iitia 0,72 2 >0, XI 14, >2^1 yl^ojt 1#tO^ >2(?:0^i Kfiyl^OJ XII 3,27 2 >0,1 >2<f«>1g0ji WlOj; *11 12,99 2 >2Ay^0j! >2<fi0^l 1<fiyl)0Jl 1970 XII 6,47 2 >0, XLtXII 12,18 2 >2<fl>1oOJ >2<f«0o >2omgj T a b e l 3 8 Biineva pullide arvu koosesinemine erineva arvu lehmadega leete s и dt г leoie*tiliseet uuremed sagedusest tumuvpli välkeenvd X 1,34 2 >0,5 1964*1965 " 0,36 2 >0, " 10,06 2 >2<f$1g; ,55 2 >0,1 1^10^1 >2/l > ,06 2 >0, , lao$ «,19 2 ^2/l>2 i >ž/j1j la>2^; 1966 или O.Ol 2 >0, XI 7,eo 2 1»69 XXX 16, >2/l>2(l l/l> MCI «.» 2 >2/lögl 1970 XII >0, МП *.** 2 >0,05 >2/1091 X fiiy t i/>0 14
.")

16 koos ühe vasikaga psdralahmaga kui vasikata EMxalS&g&Bai ülekaalus on kompositsioonid >24 г > 1y0j ja 1<f : > 1yj ning teoreetilisest harre* esinevad kompositsioonid *2<f : :>1gj, >2^ t Oy ja 1< i 1y0j. 4*3.4. Brfowra arvu рцд,щ? ^oacelnemlne erinera arm lehmadega Oktoobris on juhuslikust jaotunuseet usaldataval ta-> semel erinenud ainult 1967.» Ja a. andmed ning kokkulangevad on ainult kahe viinaee aasta analüüsi tulemused (tabel 38) a. esines sagedamini koos kaks Ja enam pulli ühe lehmaga ning harvem kaks ja enam pulli kahe vsi enama lehmaga, riimastel aastatel aga vastupidi a. novembri ja a. oktoobri+norembri kompositsioonide esinemissagedused olid sarnased a., a. detsembri andmed aga ja a. oktoobri andmetega. Hagu eespool toodud andmetest selgus, on Juhuslikust jaotusest tunduvalt erinerate kompositsioonide arv märksa väiksem esinenud kompositsioonide üldarvust. Võrreldes prevaleerivate vsi oodatust väiksema esinemissagedusega kompositsioonide esinemist aastate lslkes (tabel 39) näeme, et prevaleerivad kompositsioonid on enamail juhtudel jäänud samadeks, väiksema esinemissagedusega kompositsioonid on aga eri aastail erinenud sagedamini. Oktoobrikuul olid juhuslikust jaotusest tunduvalt erinevad ksige sagedamini järgmised kompositsioonid (sulgudes märgitud, mitmel aastal v8i aastagrupil oli väiksema e sinemi seagedusega): prevaleerivaks/ : 1^0j (6/0) >2<# s 1$ >1J (0/3) 2^1 j (5/0) 0<? :>2$OJ (1/4) 06 s 2y >2j (5/0) 06 :>2y1J (0/6) 1 & : 2jOJ (3/1) li i 2? >2j (1/5) >2i :>2y (3/1) >2<f : 0? (0/3) Novembrie-detaembrie toimuvad kompositsioonide esinemissageduetes Järgmised olulisemad muutused (tabel 39)t sagedamini esinevad koos )2<f s > 2g0j, sagedam on 1$ >2J esinemine pullita, juhuslikust jaotumusest harvem esineb 16 s 1y > 2J ning >2<? : 1g0j. 15

17 ö Ф -d r-i Ф - (JN H В кч Ю Ф > 5» Н H та со зд <D т) ф -р & еюн СО л ш и I» ä в Ф -d iz<i «in - к>ч>г< 4Г<««iii W_ L ГЙЬЙ Poll. lo«fzt Pwln. Poll«. *0» />г<»,и /о«ргсбг и fzt.». fit + /o»p^t /г<> /К» «/Ю«Р«.*Г< ^г<». /t-<««^pptcl-ži, ;Г<» > И * ^0*Р0оС< /Г<»,И»О«РОБГ< *?г*>. ри PO'FOL I- OO X XX XX X oo 00 XX 00 о 0 X XX X OO OO 00 XX XX XX XX XX XX XX X oo о X X XX XX XX XX 00 о 0 К X о 00 XX XX XX XX X oo X о о X 0 OO X о X XX X X XX XX XX XX X XX О OO 00 X 0 XX XX 00 XX XX OO XX Л Л N в öx О r- T 1- Г V 1-1- r* t * $ $ OO 0 XX X о X XX X XX XX XX OO 00 oo oo oo XX XX XX XX oooo о о oo oo $ $ $ d a4 M И H >$ H M л и


18 4.4. KOHPOSITSIOOUDE esuhcs- SAGKDÜSTB КВЯГКТОЗТВ PÕHJUSED Teatud ковров!teioonlde juhuslikust jaotusest sageda- Blnl v8i harvemini eelnemise pidevue näitab, et eri eleaentide liitumine grupiks pole juhuslik, vaid eeaduepärane. Arvestades kirjanduses toodud andmeid /15, 33, 45, 89, 91, 201/, esinevad eri elementide vahel järgmised sotsiaalsed seosed: ad.cf : ad.y - jooksuajal generatiivne aflinsua; ad. v6i subad. у i juv. - generatiivne afllneus; ad. ja eubad. t?ct - väljaspool jooksuaega afiinsus; ad. ja eubad. do" - jooksuajal tsukumine. On esitatud andmeid/45,89/, mille järgi eelmise aasta vasikaist jäävad lehamulllkad emal о oma juurde ka järgmiseks aastaks, seda isegi siis, kui emaloom toob uued vasikad. Pullmullikad aga lahkuvad tavaliselt peale esimest talve eealooma juurest. Hagu eespool nägime, on Eesti põdrapopulatsioon niinimetatud avatud populatsioon ja vaadeldaval ajavahemikul on esinenud nii immigratsioone kui ka emigratsioone, kaheldamatult on mõjutanud ka populatsiooni mis sotsiaalset struktuuri. Immigratsiooni üheks indikaatoriks on Isasloomade osa järsk suurenemine, mis vdimaldab vaadeldaval perioodil eraldada "paikse" populatsiooni ksrval "migratsiooni "-populatsioone, s. t. alasid, kus antud aastal eelneb tunduval arvul Immigrante. Nende kahe emalüüsirajoonide rühma sotsiaalse struktuuri võrdlus näitab olulisi usaldatavaid erinevusi eri komponentide esinemissageduses (tabel 4C. T a b e l 4 0 Niinimetatud "migratsiooni-" ja "paikse" populatsiooni sotsiaalse struktuuri erinevused isendilistes gruppides Kompositsiooni esinemissagedus Kompositsioonid "migratsiooni" "paikne" X 2 f popul. popul. paaria rohkem vähem 4,81 <0,05 Pullita lahiß vasixa^u (-9J) rohkem vähem 7,12 <0,01 üksikult esinevad pullid (1<4 ) rohkem vähem?,95 <0,03 UksiKUlt esinevau lehmad (-1^-) vähem rohkem?,34 4),05 >2 pulle võrreldes üksikute pullidega rohkem vähem 6,31 <0,01 Ühe lehma kohta pulli rohkem vähem 5,o2 <0,025 ühe pulli kohta 2 #.,4 lehma vähem rohkem 4,52 <0,


19 1 ф т) н р Й о а о СО о а, * Ф о со ZS аз а, и 3 4 '-0 03 а ^ ч -н d ч 0 -Р ОА СО га Ф S р С/3 ф н -з j I -н Н с d й о а н 'ta со н Р 1 л -Г\-Н ф ч о» 8г< KN r* с <м r* Гг< r* V Fl 10 FM- я\ с4и- 00 О. JC F ip - V F 4j> - ч Ю * V O 1 1 tx D TV d- K\ CM Ol СО IN <-»-аа *А «*?'<?? СО кч 1Л О г??? Ž CV Л е? е? *? N V О ^ v3 0 0 d - < M K > 5 f C M < r * r A C v i A 1Л 1Л ^ О КЧ Г C\J О КЧ 1- Ч Ч Ч Ч Ч 9 Ч Ч Ч? С^.СЧСОСЧКЧС>.0-*1Л е» « я» Н К Ч Г ч О K N I T S C M К > V D V со la U3 IN ia Cv vo " * * * н Ч Ч Ч Ч Ч Ч Ч Ч tr V СМ (Т> vd О К> г г ^1Л 00 О??? ч ч? 9 S Cv CM ra ;м 1А Cv О 00 о s> la Cv см <Т\ ко d- '???? $ < О о d- О О d- ГА О см ha чо d* о О СО d"???? о?? ч О о ГА IA СО О IN <0 CO СО V. ^ia СО???? ~ч? i гч d* г_ см со чо A CV CV о см?? 0 1.< ч < мл Cv г>- СМ Q ГА vo (N??? f < cm СО ia Cv Cv см '???? S 8 о??? г\1 м ГГх (S. И ч ч н т- гч *ч d- la ^OCs-IOCTiOv-vAirA м к


20 Lisaks eespool esitatud andmetele oli kompositsioonide esinemissageduste erinevuste põhjuste analüüsiks võimalik kasutada Mahtra Riiklikus Jahlmajandis aastal kogutud detailsemaid andmeid eriranueeliste pullide Ja lehmade koosesinemietest,andmeid eri komponentide esinemissageduste dünaamikast vaadeldavatel aastatel ning eri komponentide dünaamikate korrelatsioonide alusel selgitati nende komponentide vastastikuste seoste suund Ja tugevus (tabel 41, Joon. 32). Kõigi nende andmete põhjal vsime teha Järgmised olulisemad Järeldused. 1. PÖdragruppide kompositsioonide esinemissageduste määramisel on oluline osa vasikate esinemissagedustel, s.o. populatsiooni produktiivsusel. Sealjuures ühe vasika esinemine grupis on positiivses korrelatsioonis pulli ja lehma koosesinemisega, kahe ja enama vasika esinemine korreleerub positiivselt aga pullita kompositsioonide esinemissagedustega. Vasikate esinemissagedusega negatiivses korrelatsioonis on ainult pullidest ja ainult lehmadest koosnevate gruppide esinemissagedus, kusjuures pullidest koosnevate gruppide esinemiseagedus korreleeerub tihe ja lehmade esinemissagedus kahe ja enama vasikaga gruppide esinemissagedusega. Ühe vasikaga emaloomade Juures on eelistatud noored pullid: on võimalik, et halbade kasvutinglmuetega suve-talve Järel on need sama põd"^**"» eelmise aasta pullvasikad: Alaskas kogutud andmeil /89/ olid vasikaga põdralehmadega koos esinevad mullikad eranditult samade lehmade eelmise aasta vasikad. Kahe vasikaga emaloomadega (s. o. vanemate põdralehmadega) esineb pulle need on kas ainult noored (sama põdralehma eelmise koos harva, aasta vasikas pro vasikad) või nendega on liitunud ka üks vanem pull. Võimalik, et see on põdralehmaga liitunud juba jooksuajal. Kui kompositsioonides mitte arvestada vasikate esinemist ja piirduda ainult adulteetega, nagu seda tegi D.Houston /33/, siis ilmneb eri aastate andmete väga suur kokkulangevus (tabel 42). Võrreldes kahe viimase kompositsioone eri suurusega gruppide lõikes vaatlusaaeta abil, ei esine usaldatavaid erinevusi üksikute Ja 4-isendlliete grup-


21 Tabel isendiliste gruppide kompositaioonide esinemissagedus (protsentides vaetava suurusega grupis) oktoobris; vasikad on kompositsioonidest välja jäetud ürupi suurus komp. % _ 1 Gr. Gr. _ Gr. % Gr, Aasta t 1 BTV arv arv arv 19M в ? ü u T a b e l 4 ) Põdra eri suurusega gruppide esinemissagedus protsentides oktoobris aastail (summeeritud kogu territooriumi andmed) laeta Uruni suurua >9 Kokku grupp ,7 22,9 18,5 12,1 6,4 3, ,2 3, ,7 24,7 17,1 13, , b,3 26,3 19,6 12,4 6,9 4, , ,5 26,3 19,8 11,4 5, , O5 39,0 <=4,4 18,6 10,1 3, , ,2 25,6 17, , , ,3 25,6 17,8 10,9 3, ,4 26,3 17,9 10,9 7, , ,1 26,7 20,1 11,4 4, ,3 25,4 18,7 11,5 6, , ,6 15,7 8,4 6,6 4,1 2,

 koosnevate gruppide esinemissageduste negatiivne korrelatsioon kõigi vasikaga gruppide esinemissagedustega näitab, et tunduv osa ahtratest või vasikata jäänud lehmadest")
22 pide kompositsioonid langevad kokku 99 usaldatavusega (Я; 2, vastavalt 0,00003, df = 1 ja 0,22, df * 2). Samal ajal gruppide keskmine suurus on aastal tunduvalt suurem aastal täheldatust: üksikuid on aastal üle kahe korra, vähem, neljaliei gruppe aga ühepalju. 2. Ainult pullidest (pullist) ja lehmadest (lehmast) koosnevate gruppide esinemissagedus on negatiivses korrelatsioonis kõigi teiste komponentide esinemissagedustega. Ainult lehmadest (lehmast) koosnevate gruppide esinemissageduste negatiivne korrelatsioon kõigi vasikaga gruppide esinemissagedustega näitab, et tunduv osa ahtratest või vasikata jäänud lehmadest ^«tävad omaette, liitumata pullidega. Ainult pullidest (pullist) koosnevate gruppide esinemissageduste tugev negatiivne korrelatsioon vasikaga esinemissagedusega (r Q = -0,67 ) näitab pullide gruppide afllnsust vasikaga emaloomade suhtes: ühelt poolt esinevad noored pullid halbade kasvutinglmustega aastate Järel koos emalоomaga (vt. eelmine punkt) ja teiselt poolt heade tingimustega aastatel, mil jooksuajas osalevad ka lehnmullikad, näivad pullid eelistatult liituvat esmakordselt poeginud noorte lehmadega. 3. Kuna põdra sotsiaalset struktuuri mõjutavad populatsiooni produktiivsus ja noorte suguküpsuse saavutamise iga (sõltuvalt suvistest-talvietest tingimustest), ei tohi eri aastate andmeid liita eelneva statistilise kontrollita. Sama kehtib ka eri kuude andmete liitmise kohta, sest eri aastail võivad agregeerumiet põhjustavad tingimused ilmneda eri kuudel. Kahjuks käesoleva ajani sellest pole juhindutud, mis teeb võimatuks eri aladelt pärinevate andmete võrdlemise. KSik eeltoodud andmed näitavad, et põdra normaalses populatsioonis tüüpiliste emaperekondade kõrval esinevad püsivalt ja teatud arvulietes suhetes ka teistsugused sotsiaalsed grupeeringud. Sellepärast pole ka Õige nimetada põtra tüüpiliseks üksi või ainult ema perekondadena elavaks liigiks. PÕdragruppide kompositsiooni analüüsi praktilist kasutamist võib ette näha 1) populatsiooni produktiivsuse hindamisel (indikaatoriks ühesooliste gruppide väiksem või suurem suhteline esinemissagedus), 2) populatsioonis 21 ahtrate

23 emasloomade suhtelise eeinenieeageduee hindamisel, 3) populatsiooni vanuselises struktuuris toimuvate muutuste selgitamiseks. Erisuurusel!ste gruppide esinemissageduse sõltumine populatsiooni sotsiaalsest struktuurist võimaldab suhteliselt kergesti kogutavald andmeid kar.lade suurusest kasutada sotsiaalse struktuuri Indikaatorina ning samal a.1al näitab frekvents! sobimatust populatsiooni arvukuse hindamisel.
 antud liigi erineva asustustihedusega levik talle elutingimustelt sobival alal; seda vsib omakorda vaadelda (1.")

24 5. POPULATSIOONI TERRITORIAALNE STRUKTUUR üheks populatsiooni struktuuridest on ka populatsiooni territoriaalne struktuur, в. о. antud populatsiooni soodustavate Isendite paiknemine ruumis. Territoriaalset в traktuuri võime vaadelda kolmel tasemel: (1) antud liigi erineva asustustihedusega levik talle elutingimustelt sobival alal; seda vsib omakorda vaadelda (1.1) liigi levikuna antud momendil ja (1.2) liigile elutingimustelt sobiva ala levikuna, mis määrab liigi võimaliku (perspektiivse) leviku; (2) liigi levik erinevates elupaikades, s. o. liigi elupaigaline ehk blotoobiline levik, mida S.S. Schwarz /216/ on nimetatud ka populatsiooni topograafiliseks struktuuriks: (3) liigile iseloomulikust eluviisist sõltuvalt kas karjade, perekondlike gruppide v8i tiksild.eendite levik liigi poolt asustatud alal. Kui esimesel kahel tasandil populatsiooni territoriaalse struktuuri vaatlemine on üldlevinud, siis kolmandal tasandil, mis puudutab populatsiooni moodustavate isendite reaalselt koos esinevaid, elutsevaid ja koos tegutsevaid grupeeringuid, on populatsiooni territoriaalse struktuuri vaatlemine rakendamist leidnud suhteliselt harva ja peamiselt kas tüüpiliselt karjallse eluviisiga ssraliste v8i koloniaalse eluviisiga näriliste puhul. Indiviidi (isendi) lugemine populatsiooni väikseimaks ühikuks on 8ige ainult teatud, aga mitte ksigil juhtudel. FShimasB isenditest pole looduses levinud mitte üksikutena, vaid erineva isendite arvuga gruppidena. Nii on see ksigil aastaaegadel, kuna nende järglaste areng on suhteliselt aeglane, mis tingib emaperekondade aastaringset esinemist ning indiviidide vahel esinevad ka teised seosed, mis tingivad vähema v8i suurema arvu isendite koosesinemist. Seega populatsiooni territoriaalse leviku vaatlemise piirdumisel ainult kahe esimese tasandiga, iseloomustades levikut esj.nemlse-puudumiae ja erineva asustustiheduse kaudu (1 s e n - d e 1 d pindalaühiku - enamasti 1000ha - kohta), on tegu 23
 kui ka teoreetiliste küsimuste lahendamisel. Tuntuim põdra ja metsa vahekorda uuriv spetsialist A.A. Kozlovskl /138/ väidab otee, et \".")

25 looduse в eel kujul mitteesineva abstraktsiooniga, mille realiteediks pidamine võib kaasa tuua olulisi möödalaskmisi ja vigu nii praktiliste-metoodiliste (näit. loendusmeetodid) kui ka teoreetiliste küsimuste lahendamisel. Tuntuim põdra ja metsa vahekorda uuriv spetsialist A.A. Kozlovskl /138/ väidab otee, et "... aeustustihedus ei iseloomusta tegelikku kohapealee põdrapopulatsiooni jaotumuet. Aeustustiheduse kasutamist piiratud territooriumidel sõrallste leviku iaeloomustamiseke ei pea võimalikuks ka A. Bubenik /9/. Väga teravalt tõstatas küsimuse liigee generaliseerimiae kahjulikkusest ja vajadusest tundma õppida populatsioonide konkreetset, tegelikku jaotumust P.B. Jürgenson oma aastal ilmunud artikliа /230/. Eriti rõhutas ta populatsiooni territoriaalse struktuuri tundmaõppimise olulisust seoses uute loenduameetodite väljatöötamise vajadusega: "Viimastel aastatel taandatakse ulukite loenduse ja selle meetodite probleem praktiliselt arvude seeriatele ja variateiooniatatistikale tulemuste tõenäosuse hindamiseks. Tegelikult on see probleem looduslikest seaduspärasustest loesdusühlkute (sõrendus minu, H.L.) paiknemisest ruumis ökoloogilisel ja etoloogilisel alusel." poolt metsale tekitatavate kahjustuste vältimise Ka sõrallste abinõude väljatöötamise üheks eelduseks peab P.B. Jürgenson (nagu ka A. Bubenik /8, 104/) teadmisi populatsiooni konkreetsest territoriaalsest levikust, nimetades seda "... mitte vähem olulieeks momendiks, kui selle populatsiooni toidureseurseide paiknemist." Oma postuumselt ilmunud raamatus /^32/, käsitledes populatsiooni struktuuri uurimist, nimetab ta kahte põhisuunda: 1) levinumaks on piirdumine vanuselise ja soolise struktuuriga, seostamata aeda konkreetse territoriaalse analüüsiga, ning abstraktsete keskmiste "karjaeuee" koefitsientide ja nende sesoonsete muutuste uurimine; 2) "... uuritakse reaalselt looduses eksisteerivaid populatsiooni struktuurseid grupeeringuid ja nende dünaamikat kindla territooriumi piires, teisiti öeldes - uuritakse territooriumil erinevalt paiknevaid struktuurseid grupeeringuid... Taolist laadi uurimused, millel on nii ökoloogiline kui ka etoloogiline iseloom, hõlmavad ka struktuuri põhjuste uurimist ning praktilisi järeldusi populatsioonile." Вeimeeee suunas teostatavaid uuringuid peab ta praktikas kasutamiseks mitteküllaldasteks. 24


26 SSraliete territoriaalse struktuuri vaatlemisel oa kahtlemata populate!ooni väikseimaks ühikuks enamail juhtudel isendite grupp ja harvem flkeikisead«et mitte rääkida ükeik-grupiliseet esinemisest, on järgnevalt populatsiooni moodustavate isendite jaotumust käsitledes loetud omaette grupiks ka üksikisendi territooriumi kasutamisel ja levikul ruumis pole sisulist erinevust üksüelaendi v8i kahe- jne. isendilise grupi vahel - ksik nad territooriumi kasutavateks primaarseteks»ы kutaka. Populatsiooni territoriaalsel analüüsil grupilise acalüüsi harval kasutamisel on ka oma objektiivsed põhjused: kas loendatakse indiviide ning mitte gruppe ja neis olevaid indiviide, loendusmeetod võimaldab hinnata ainult on suhtelist esinemissagedust (loendusjälgede, ekskrementide alusel jt.), v8i saadakse materjal suhteliselt väikesearvulisena ja pika aja vältel, mis ei vsimalda saada usaldatavaid andmeid ühe kindla perioodi kohta, sest gruppide suurus muutub seaduspäraselt, mis ei luba liita eri kuudel kogutud andmeid. Grupp pole vaadeldavail liikidel p5hiühikuks mitte ainult populatsiooni territoriaalses, vaid ka sotsiaalses struktuuris: sotsiaalsed, käitumusllkud, etoloogillsed isenditevahellsed suhted määravadki grupi koostise - mis sugu, kui vanad ja millises sugulussuhtes Isendid sinna vslvad kuuluda - ning selle kaudu ka grupi suuruse. Seega vsib eo ipso arvata, et juba grupi suurus - täpsemalt: eri suurusega gruppide esinemissagedus - vsib anda informatsiooni populatsiooni soolise, vanuselise struktuuri ning produktiivsuse kohta; andmete saamine gruppide suurusest on aga vsrratult lihtsam kui andmete saamine iga indiviidi soost vanusest. Erinevused eri alade eri suurusega gruppide ja esinemissagedustes v8i nende ajalised muutused vsivad anda informatsiooni muutustest populatsiooni soolises ja vanuselises struktuuris vsi populatsiooni produktiiveuses. Käesoleva analüüsi eesmärgiks on selgitada a) jaotumuse* muutlikkus sügis-talvisel perioodil,territoriaalne muut- Siin ja edaspidi kasutatakse T. Prey poolt P. Greig- Smithi /25/ tõlkimisel kasutatud terminoloogiat: jaotus - matemaatiline, jaotumus - looduslike objektide jaotumusviis. 4 cl S
 22,4 18,0 12,9 9,1 5,2 2,6 0,4 3,0 232 1963 27,7 20,3 24,0 13,3 7,0 2,7 2,3 0,7 2,0 300 19b4")

27 T a b a l 4 Л Põdra eil suurusega gruppide esinemissagedus protsentides novembris aastail (summeeritud kogu territooriumi andmed) *..u Qrupl iuuim iokku »9 grupp«1эы 20,9 21,В 23,2 11,8 9,5 5,7 3,6 2,4 0, ,) 22,4 18,0 12,9 9,1 5,2 2,6 0,4 3, ,7 20,3 24,0 13,3 7,0 2,7 2,3 0,7 2, b4 21,5 26,3 22,3 12,4 8,4 2,2 2,2 1,1 0, ,4 22,7 20,9 9,3 5,8 7,0 5,8 1,7 2, ,6 27,8 16,5 17,4 5,2 4,3 1,7 2,6 1, ,3 22,9 24,0 1,0 4,2 4,2 4,2 3,1 5, , ,6 11,8 7,5 4,9 2,6 1,6 1,0 30b ,4 23,5 23,7 12,1 8,1 5,3 3,7 0,8 2, ,0 21,9 19,9 12,0 8,7 3,7 6,2 2,2 4, ,6 21,* 16,8 16,2 11,0 8,0 3,0 2,2 7, T a b e l 4 5 Põdra eri suurusega gruppide esinemissagedus protsentides detsembris aastail (summeeritud kogu territooriumi andmed) tirupi suurus Kokku - aata 1 i >9 gruppe 19Ы 13, 4 11,5 26,4 18,4 10,3 3,4 4,6 1,1 5, Õ2 18,6 22,0 25,4 13,6 5,1 3,4 10,2-1, ,9 17,9 14,7 11,6 15,8 3,2 4,2 4,2 10, ,8 14,9 13,9 8,9 7,9 4, ,9 16,4 24,6 14,2 6,0 8,2 3,7 3,7 2, oo 27,8 28,9 20,0 10,0 16,0 1,1-2, ,о 33,7 19,6 7,6 3,3 4,3 3,3 3,3 5, job 20,7 27,4 19,4 10,5 8,4 3,0 4,6 3,0 3, ,7 25,9 19,9 12,7 8,2 3,0 3,2 1,6 2,8 4Э ,0 22,8 20,7 14,5 7,6 5,3 5,3 1,8 3, ,0 24,1 19,6 15,3 8,8 5,5 5,1 2,8 4,7 613 $
 kogu analüüsi alusel teha metoodilised järeldused jaotumuat iseloomustavate andmete käsitlemise ja analüüsi viisidest.")

28 likkus ja muutlikkus eri aastatel, b) võimalikud jaotumuettittbld ja nende esinemise seaduspärasused, c)jaotumust määravad tingimused, d) millist informatsiooni võib saada jaotumnseet populatsiooni kohta ning e) kogu analüüsi alusel teha metoodilised järeldused jaotumuat iseloomustavate andmete käsitlemise ja analüüsi viisidest. Sendele küsimustele vastuste saamisel on võimalik analüüsida jaotumuse kui ühe populatsiooni territoriaalse struktuuri komponendi seost teiste populatsiooni struktuuridega - vanuselise, soolise ja sotsiaalsega. Analüüsil olid kasutada andmed grupi kohta, nelert on oktoobris kohatud 7811, novembris 3758 ja detsembris 2628 gruppi. Bri suurusega gruppide esinemissagedus kuude Ja aastate l8ikes on esitatud tabelites 43, 44 ja 45. Jaotumuste eamasuse-erinevuse selgitamiseks kasutatakse peamiselt X- Ja ^-meetodeid /4, 16, 186/. Kolmogorovi-Smirnovi X-kriteerium kahe Jaotuse hindamiseks põhineb nende Jaotumuste maksimaalsel erinevusel ning võrdse Jaotuaklasside Ja gruppide arvu korral näitab maksimaalse erinevuse suhet ruutjuurega gruppide koguarvust. Selles peitub kriteeriumi lihtsus ja ka põhipuudus: arvestades ainult maksimaalset erinevust, Jätab ta kõrvale kõik väiksemad erinevused, mistõttu see informatsioon Jaotumuste erinevustest läheb kaduma. Eriti suur informatsiooni kadu esineb mitmetipuliste jaotumuste võrdlemisel. Informatsiooni kaotusest on vaba Pearsoni ^-meetod, mis võtab arvesse kõigi Jaotueklasside erinevused. Sel põhjusel ongi erinevate Jaotumuste võrdlemisel ja Jaotumuste eamaeuse-erlnevuse hindamisel alljärgnevalt kasutatud ^-kriteeriumi. Kuna eri jaotumised on erineva klasside ja järelikult ka erineva vabadusastmete arvuga, ei saa jaotumuste sama- 2 sust-erinevust hinnata otse 3^ väärtuste Järgi, küll kõlbab selleks aga vastavalt igale vabadusastmete arvule määrav esinemise tõenäosus P (%-des). See näitab nii samasuse kui ka erinevuse usaldatavuse määra: jaotumuste erinevus on loetud usaldatavaks, kui P < 5 %. väärtuste esinemise tõenäosuse hinnangu P kasutamine annab Jaotuste samasuse-erlnevuse kvantitatiivseks võimaluse hindamiseks ning kõigi aastate jaotumuste võrdlemiseks graafilisel kujul.


29 P'X 0.1т h-~ го Joon. 3j>. Põdrakarjade jaotumuste sarnasus-erinevus naaberkuudel jalusel, pidev joon - november võrreldes oktoobriga; katkendjoon - detsember võrreldes novembriga; ringid - detsember võrreldes oktoobriga. j ооп 'õdrakarjade jaotumuste sarnasus-er ingvus naabaraastate oktoobri (pidev joon) ja detsembri (Katkendjoon) kuuue vahel X-alusel.
 v8i mitte. 5.1.")

30 Real juhtudel oa kasutatud ka 2 aditlivauee reeglit, ais lubab vsrreldavate Jaotumuste ühtlikkuse hindami- 2 seks liita k8igi jaotumuste vahelised v~d ja easuti vastavate vabadueastnete väärtused! vaetava 3^ esinemise t8enäoeua P vslmaldab otsustada, kas vsrreldud jaotumuai. afceeldavat kogunikku v8ib käsitleda tervikuna (ksiki jaotumus! liidetuna) v8i mitte JAOTUVUS SÜGIS TALVISEL PERIOODIL 5.1.1«Eri kuude jaotumuste ygrdlua On üldtuntud, et psdra karjaeue aasta jooksul pole püsiv, vaid muutub kaunis suurtee piirides /32, 182, 202, 211, 236/. See ei luba ka ühest kohast, kuid eri kuudel kogutud andmeid ilma nende eelneva analüüsita ühe kogumina vaadelda. Eriti järsud muutused jaotumuses toimuvat talve saabumisel, millal algab suuremate karjade moodustumine. Kuna Eestist pärinevad andmed põhiliselt eügie-talve kohta, eriti vajalik selgitada oktoobri-novembri ja novembri-deteembri jaotumuste vbrdlemisega nende samasue-erinevus. Kasutades samasuee-erlnevuse usaldatavuse kriteeriumina eelnemise t&enäoeust P protsentides, on joon.33 esitatud sama aasta naaberkuude jaotumuste samasus-erinevus. Eri kuude jaotumuste sarnasuse alusel (tabel 46) vsib vaatlueperioodi aastad jaotada kolme rühma, mida alljärgnevalt analüüsime lähemalt. I. Haaberkuudel oluliselt mitteerinevate jaotumustega aastad vsib omakorda jaotada kahte alarühma selle alusel, kae ka oktoobrikuu jaotumus detsembrikuu jaotumusest oluliselt erineb v8i ei erine. Lähemal vaatlusel selgub, et ksieil neil aastail - ka ja toimusid samasuunalised.jaotumuse muutused (Üksikute isendite arvu vähenemine ja suurema isendite arvuga gruppide on osatähtsuse suurenemine oktoobriga võrreldes), kuid need muutused olid suhteliselt väikesed võrreldes teistel aastatel muutustega. toimunud
 1962 1961 1963 1965 1966 1964 1968 1967 1969.")
 ^^\" eei^^jqi8tluekuud Ja gruppide euuiused XII KllaaatlUeed tingis used 1 2-3 4-5 >6 1 2-3 4-5 >6 1 2-2 >G X XI X keelaniete fc»usma -13-7 +56-8 -20 +2t> XI -18 XII 21-29 X ebeol.")
31 T a b e l 4 6 Põdragruppide eri kuude jaotumuste sarnasuse-erinevuse alusel eristatud aastate rühmad 16 II III Oluliselt ei erine I - XI; XI - XII Oluliselt ei erine ice X - XII Oluliselt erineb X - XII Oluliselt ei Oluliselt ei erine X-XI erine XI-XII (erineb XI-XID (erineb X-XI) T a b e l 4 7 Põdra eri suurusega gruppide protsentuaalse esinemissageduse korrelatsioon klimaatiliste tingimustega (r x 100, N=45, rajoonide ja kuude lõikes aastail ) ^^" eei^^jqi8tluekuud Ja gruppide euuiused XII KllaaatlUeed tingis used > > >G X XI X keelaniete fc»usma t> XI -18 XII X ebeol. «talau-t* а urnina -$$ IB -25 *20 XI -11 XII 14 XI luelkattega päevi 27 И5 * *24 -И9 XII " >7 *6 Talve liuneügavuste auna *28 * $1 +15 <16 30


32 II. 1963«aastal novembrikuu jaotumus ei erine oluliselt oktoobrikuu jaotumusest ning hüppeline jaotuse muutue toimub alles detsembris. Siiski ilmneb juba novembris eriti 2-ieendiliste karjade vähenemine Ja 3- ja üle 7 ieendlliete karjade osa suurenemise tendents; detsembris suureneb aga järsult üle 5 leendlllete gruppide osa (umbee kaks korda rohkem kui novembrist). III. Viiel aastal täheldati jaotumuse hüppelist muutumist novembris, kusjuures detsembris jaotumus oluliselt enam ei muutunud. Kõigil neil aastail vähenes novembris oluliselt 1- ja 2-ieendiliste karjade osa ning suurenes 3- ja rohkema isendiliste karjade osa (ainult aastal vähenes ka neljaste karjade osa)» Detsembris vsrreldes novembriga Ja aastal Jätkub väiksemate gruppide osa vähenemine ja neljaste ning suuremate gruppide osa suurenemine, 1969., ja aastal aga esineb märgatav anomaalial neil aastail väheneb märgatavalt viieste Ja suuremate gruppide osa ning suureneb 2- kuni 4-ieendillete gruppide osa. V8lb arvata, et see anomaalia on pshjustatud neil aastail rakendatud suhteliselt suurest laskenormlstt suuremate gruppide lagunemist vsis põhjustada nii osa isendite küttimine kui ka pidev häirimine. Jaotumuse oluline muutumine novembris langeb Ja aastal kokku madalate absoluutsete temperatuuride Ja aastal ka varase lumi katta tekkimisega (kolmes rajoonis lumekattega päevi novembris 20). Pftdra karjaaust määravate tingimuste selgitamiseks arvutati korrelatsioonid eri suurusega karjade esinemissageduste ja rea klimaatiliste tingimuste vahel aastatel , millal küttimise segav m8ju oli veel väike (tabel 47). Suhteliselt nsrk on oktoobrikuu keskmise temperatuuri ja karjade suuruse vaheline seos: külma oktoobri puhul (keskmiste t -de summa väikseim!) on rohkem suuremaid gruppe oktoobris ja novembris, novembris on oluliselt vähem isendilisi gruppe. Novembris ja detsembris seos samasuunaline. Oluliselt pole tugevam on absoluutsete miinimumtemperatuuride msju - oktoobrikulmade puhul suureneb isendiliste gruppide osa, mõju Järgmiste kuude karjasuaele pole usaldatav. Loogiline seos esineb lumikat- 31


33 fcega päevade arvu ja lumesügavuste summa ning Icarjaeuae vahel, kuigi seose tugevus pole enaelkul juhtudel usaldataval tasemel: seose suund langeb aga (ühe erandiga) ühte - suu- rftmn inmivattbpa gäevade arvu.1a suurema lume sügavusega talvedel on 4-.la зр^ыгяп я1 gendlllste kar.lade osa suurem. F8drakarjade suunise jaotumuse dünaamika kohta sügistalvisel perioodil vsib eelnenu alusel teha Järgmised järeldused. 1. Võrreldes eri kuude jaotuauei Ja nende dünaamikat, vsib eristada kaht pshillet jaotumusttiüpl: sügisene ja talvine. Sügisest jaotumustüüpl iseloomustab suhteliselt suur 1- ja 2-isendiliete gruppide esinemine, talvist jaotumustüüpl aga suurem 4- ja enamalsendil!ste gruppide osa ning väga suurte gruppide suhteliselt suurem esinemlssagedus; v9rreldee sügisese Jaotumuatüübiga on talvises üksikuid яппяап aastail ainult , üksikute ja kahellste gruppide summaarne osa on aga % sügiseses jaotumustüübis esinevast. 2. Kõigil aastail on novembris v8rreldee oktoobrikuuga toimunud 1- ja 2-isendiliete gruppide arvu suhteline vähenemine ning suuremate gruppide osatähtsuse suurenemine. Kuuel aastal Üheteistkümnest pole siiski novembrikuu jaotumuse erinevus küündinud statistiliselt usaldatavale tasemele (eriti väike oli erinevus ja aastal), vilel aastal aga tolmus järsk sügiseselt jaotumustüübilt talvisele üleminek juba novembris - ilmselt seoses varase külmaperioodi v8i lumikatte kujunemisega. 3. Hüppeline talvisele jaotumustüübile üleminek detsembrikuul on toimunud vaadeldaval ajavahemikul ainult aastal. Pikaldane järskude muutusteta talvisele jaotumustläibile üleminek on esinenud neljal aastal (detsembriks saavutati siiski oktoobrikuu jaotumusest statistiliselt usaldatav erinevus), ja aastal tõenäoliselt Jaotumuetüüpi detsembrikuuks aga ei saavutatudki. talvist 4. Eelnenust tuleneb metoodiline järeldus: jaotumuste erinevusi eelnevalt kontrollimata ei tohi liita (koos käsitleda) eri kuude jaotumuste andmeid. Et samasuunaline jaotumuse muutumise tendents esineb aga igal aastal (kuigi sa- 32


34 gell statistiliselt mitteolulisel taeemel), 011 otstarbeka a võrrelda sügisese jaotueuettmbl analüüsil ainult oktoobrikuu Ja talvise Jaotumustüübi analüüsil ainult detsembrikuu Jaotumas! Jaotumus oktoobris Võimaliku kindlasuunalise Jaotumuse muutuse selgitamiseks, mis võib olla tingitud pidevast populatsiooni arvukuse suurenemisest, analüüsiti naaberaastate Jaotumuste samasust-erinevust (joon. 34). Hagu võib näha oktoobrikuu andmete võrdlusest, ei erinenud vaatlueperloodi esimesel neljal aastal naaberaastate Jaotumised teineteisest oluliselt (kuigi esineb erinevuste pideva suurenemise tendents!), kuid viimasel kuuel aastal pole ühelgi juhul naaberaastate jaotumised olnud samastatavad: ja aasta Jaotumuste erinevus on usaldatav rohkem kui 99,95 % taeemel Й^ОЗ>78"*, df - 7), veelgi olulisem on erinevus ja aasta jaotumuste vahel (3^2 = 62,64***, df» 8) ja ainult ja aasta jaotumuste erinevuste usaldatavus on 97,5 * tasemel Ц 2-15,15 х, df - 6). Kuigi naaberaastate Jaotumised on vaatlueperioodi teisel poolel olnud oluliselt erinevad, ei tähenda see ometi, et nende hulgas ei võiks olla nii omavahel kui ka esimese poole aastate jaotumustele perioodi sarnaste jaotumustega aastaid. Selle kontrollimiseks võrreldi kõigi aastate oktoobrikuude jaotumus! omavahel 1? abil (Joon. 35). Esitatud andmeil jaotuvad vaadeldava perioodi Jaotumuste oluliste erinevuste alusel kolme rühma: 1) aastad , 1966, ; 2) aastad 1965 Ja 1967; 3) aasta. Kui teise rühma kuuluv aasta jaotumus on aastad veel nõrgalt seostatav esimesse rühma kuuluva aasta jaotumusega, siis aasta Jaotumus on täiesti erinev kõigi teiste aastate jaotumustest väga olulisel taeemel (X, 2 on teiste aastate jaotumustega (kõigil juhtudel df* 8): 19б1 т ,77*^, ,92***, ,97***, ,64***). Hende kolme aastaterühma jaotumus! võib nlmeta- 5 33

35 1969 /963 P-X P>90 90 >p? 70 70>p>W 4fl>p>26 20>p> P-X O.I<p<t I<p< Joon. 35- Sri aastate oktoobrikuu põdrakarjane jaotumuste sarnasus-erinevus alusel: A - sarnasus, В - erinevus. de sügisese jaotustüübi alatüüpideks, millede erinevused seisnevad põhiliselt üksikute ning nelja-viie ja suurema isendite arvuga gruppide erinevas eeinemiseagedusee. Tuleb rõhutada, et esiletoodu earnasused-erinevused põhinevad kogu territooriumi koondandmeil, mis ei arvesta eri piirkondades esineda vsivaid erinevusi. Sende koondandmete analüüsi alusel võib väita, et samasugune jaotumus iseloomustab nii vaadeldava perioodi esimesi ( ) kui ka mitmeid viimaseid aastaid (1968, 1970) - nende aastate jaotumused langevad kokku väga suure tõenäosusega (summaarne Ц 2» 15,48, df = 24; F > 90 %). Kuna samal ajal on 34

36 aga põtrade arvukus meie alal mitmekordistunud, võime teha Järelduse, et kogu terrhtnn^iiai hõlmavate andmete analüüsi alusel erieuuruaeliste oõdrakariade esinemissagedus (jaotumus) oktoobrikuul ei sõltu põtrade asustustihedusest » Jaotumus detsembris Analoogselt oktoobrikuu andmete analüüsile võrreldi ka detsembrikuu jaotumuste eamasust-erinevust vaatlusperloodi aastate vahel. Andmed eri aastate või aastategruppide jaotumuetest ja nende jaotumuste vahelistest seostest on esitatud joom. 36 (naaberaastate jaotumuai analüüsiti eelmises osas ning andmed selle kohta olid esitatud Joon. 34) I96J P-% s==s«p> sq ===== 90>p>70 70 >/>>*» ьо>р>го I0>p> 5 Joon. 3b. Sri aastate detsembrikuu põdrakarjade jaotumuste sarnasus-erinevus X alusel: А - sarnasus, ti - erine 35

37 * * ««*

.")
38 Andmeist selgub, et samuti leul oktoobrikuu jaotumuste puhul, pole ka detsembris registreeritud karjade Jaotunrueed kõigil aaetail earaaeed. Omavahel oluliselt mitteerlnevad on a. jaotumised (samuti kui oktoobrikuu jaotumnsedkl) ning a. jaotumueed: eaimeae aastate grupi Jaotumuste kokkulangevuse tõenäosus on 40 ja 30 % rahel (summaarne = 18,46, df - 17), viimasel aastategrupil aga 90 Ja 80 * vahel (summaarne 3^2» 11,07, df - 18).Nende mõlemate aastategruppide jaotumueed gruppide koondandmete alusel on oluliselt erinevad (^2 = 20,7**, df - 8). Võrreldes vaadeldava perioodi oktoobrikuude Jaotumuste omavahelisi kokkulangevusi detsembrikuu jaotumuste omavaheliste kokkulangevustega võime nentida, et enamail juhtudel oktoobrikuu jaotumuste kokkulangevuse puhul samade aastate detsembrikuude jaotumueed usaldatavalt kokkulangevad ei ole. Järelikult kui oktoobrikuu Jaotumuste samasuse puhul võivad detsembrikuu Jaotumueed oluliselt erineda, peavad eri aastail detsembrikuu.laotumast kujundavad protsessid kulgema erinevalt (karjaeuert mõjutavad klimaatilised tingimused) või mõjutavad seda eri aastail erinev* intensiivsusega toimivad tegurid (näiteks küttimine, sisse- ja väljarändeco Woobflkuu JaiytiMiata terrf tqrtmflnn BfwitUfryiff Aastate võrdlusel opereerlsime kogu vabariigi territooriumilt kogutud summaarsete andmetega. Selgitamist aga vajab, kas Igal aastal jaotumueed vabariigi eri oeedee olid samased või erinevad, s. t. kas jaotumust määravad protsessid toimivad samal aastal kogu territooriumil ühesuguselt või eri osades erinevalt. Selleks võrreldi analüüeirajoonide jaotumusl omavahel igal aastal ning hinnati nende saua suet-erinevust Д 2 esinemise tõenäosuse alusel (joon. 37). ^2 aditiivsuse reegel võimaldab iga aastat hinnata ka.jaotumuste ühtlikkuse seisukohalt, s. t. kas antud aasta eri rajoonide jaotumus! võib lugeda kuuluvaks ühte kollektiivi või tuleb neid vaadelda eraldi. Selleks liideti kõikide rajoonide vaheliste jaotumuste samasuse erinevuse JJ 2 hinnangud (igal aastal 10, kokku 110 võrreldud paari) ja vasteeral* 37

 1 H d У i e rajoonid» 6 & I ЯгInetused usaldatavad р väärtus % 1*2 р väärtus Л 1*2 >90 >Р0 >40 >20» I* 2 $uf Р>У^,Э >99 >95 X«2 Р> ' i - В 1 5 7 10 45,39 46 <50 1 - - 29,46 9 <0,1 i")
39 T a b e l 4 8 Vaatlusaastate oktoobrikuu jaotumuste Ohtlikkus rajoonide omavaheliste võrdluste summaareeil andmeil (võrreldud kõiki rajgone, 0^2 summeeritud) leete it 2 41 J* , »>90 1Э62 42, *9, , >p» , ,1>p>0, , >p> ,26 41 P<0, , >P> ,87 56 P< ,18 76 p<0, ,48 76»«0.1 t a b e l 4 9 Oktoobrikuu eri rajoonide põdrakarjade jaotumuste sarnaa us te-erinevus te esinemissagedus ja nende usaldatavus aastail (Kaelusel) 1 H d У i e rajoonid» 6 & I ЯгInetused usaldatavad р väärtus % 1*2 р väärtus Л 1*2 >90 >Р0 >40 >20» I* 2 $uf Р>У^,Э >99 >95 X«2 Р> ' i - В ,39 46 < ,46 9 <0,1 i - С ,44 26 < ,17 29 <0,05 А - D , в 111,22 а <0,05 А - Ж ,74 31 < ,72 18 <0,05 В -С в 10 55,56 57 " ,69 5 <5 1 - В ,13 31 < ,44 21 <0,05 В * ,19 44 < ,47 12 *0,05 С -В ,79 50 < в , ,67 8 <5 D - Ж ,13 36 < ,09 8 <2,5 38
 ning 1966. Ja 1968.")

40 vabadusastmete arvule selgitati antud if väärtuse eeineni ee t6enäoeue, ale ühtlasi on antud aasta jaotuauete ühtlikkuse tõenäosuse hinnanguks (tabel 48). Selgub, et ühest kollektiivist pärit andmetena v8ib vaadelda vaatlueperioodi esimese nelja aasta ( ) ning Ja aasta jaotuausi, teistel aastatel on eri rajoonide Jaotumuste kuulumine samasse kollektiivi väga väikese tõenäosusega: 1969» aastal on eee usaldatav 1:20, ülejäänud aastatel isegi vähem kui 1:999 vastu. Detailseaal analüüsi1, mille andmed on esitatud Joon. 37 Ja 38, selgub, et rajoonidevaheliste Jaotumuste olulisi erinevusi ei sedastatud kahel aastal, kolmel aastal esines erinevusi ainult kahe rajooni ning ühel aastal kolme rajooni vahel, viiel aastal eelnes olulisi erinevusi aga enam. üheteistkümne vaatlusaasta jooksul esinenud oluliselt mitteelunevate Jaotumuste kokkulangemiete arvu Ja nende eaaasuee usaldatavuse vahel vbib välja tuua omavahel kõige "sugulaslikuna" jaotumusega Ja omavahel kõige sagedanini Jaotumuse poolest erinevad rajoonid (tabel 49)«Jaotumuste sarnasuse poolest jaguneb Eesti territoorium nagu kahte ossa: ühel pool Lääne- Ja PShja-Eesti (rajoonid A Ja B) ning teisel pool Kesk-, Ida- Ja L8una-Besti (rajoonid C, D ja e) JAOTÜMÖSTÖÖBID Oktoobrikuu.jaotumuste vsrdlus analüüsirajoonlde lslkes Olles selgitanud rajoonide andmete kokkulangevuae-erinevuse k8lgi vaatlusaastate lbikes, on nüüd vclmalik ka selgitada, kuiv8rd need jaotumueed korduvad, s. t. kas v8ime rääkida teatud Jaotumustüüpidest v8i on tegu muutustega, mis vaadeldaval juhuslike perioodil seaduspäraselt ei kordu. Sarnasuse-erinevuse hindamiseks kasutati jällegi meetodit ühe aasta omavahel mitteerinevate rajoonide summaarsete andmete (joon. 39) v8rdlemiseks teise aasta omavahel mitteerinevate rajoonide'grupi andmetega. Selline vsrdlus (kokku vsrreldi 67 paari) v8imaldas selgitada viis omavahel oluliselt erinevat Jaotumuetüüpi, millede karakteris- ЗУ

41 г - IMI - ж? 50 jf 30 ю В.CX о я зо я т я я j» /е V л.а.с о.г Я А. В. С ОС 30 ю ms ю т я 4.0 с. е.с 30- л.в С. Г я 1971 I I г 3 * $ S 7 I»Эк Joon. 38. Põdrakarjade Jaotumus oktoobrikuul aastatel usaldatavalt erinevate ana luus ira j oon lae vsi nende gruppide lõikes. 40


42 I Z 3 t >3 ie/gr. Joon. 39. Põdrakarjade jaotumus tüübid oktoobrikuu andmetel. Joonise paremal serval on toodud eri suurusega karjade esinemissageduste erinevuste amplituudid ü П I П П и и E II П * 1 и п I п я п и д Е д л I 7 л д I д д ? I ш Щ5 1 И ЩИ П.Ш 1 п! Е Д Я,? д I Щ 5,5 дш Я и.ш Е Iilil 1.Ш Е д Joon. 40. Põdrakarjade oktoobrikuu j ao tum us tüüpide (I...V) territoriaalne levik aastail 19M

 kuu X XI XII Grupi suurus 1 2.. 4....5 6 1 de 4.. 5 b 1 2....5.\".5 Af f.")
43 t a b e l 5 0 Põdra eri suurusega gruppide esinemissagedus protsentides oktoobris eri jao turn us tüüpides Jaot l- Grupi suurus üruppida mus tüüp arv >9 52,4 23,2 13,8 4,5 2, ,2 0,5 1,2 327 XI 29,9 25,5 19,8 12,5 5,0 3,4 1,5 1,0 1, III 27,6 27,3 17,8 10,4 8,1 3,7 2,0 1,4 1, IV 12,6 26,3 17,9 15,5 11,7 6,1 3,5 2,9 3,4 484 V 26,0 21,8 15,1 13,3 8,1 6,1 3,8 2,0 3, t a b a l 5 1 Mõnede sotsiaalsete grupeeringute ja sigimisnäitajäte korrelatsioon eri suuruseliste karjade esinemissagedusega aastail 1ЭМ kuude lõikes (r x 100, N=45) kuu X XI XII Grupi suurus de b ".5 Af f j juv > I51-3b И * b 2j +JJ ' ; J JJ* j JJ / j 25 JUV / oo -55-1b b B 18 42


44 tikad on esitatud joon. 39 ja tabelis 50 ning ajaline ja territoriaalne levik joon. 40. Alljärgnevalt iseloomustame eraldatud jaotumustüüpe ja nende esinemist vaadeldaval perioodil lähemalt. I j a o t u m u e t ü ü p. I s e l o o m u l i k u k s o n e r a kordselt suur üksikisendite esinemise sagedus: üle nelja korra rohkem kui IV tüübil ja ligi kaks korda rohkem kui teistel ülejäänutel. Võrreldes kõige sagedamini esinenud "normaalse" II tüübiga, on erinevad ka teise suurusega gruppide omavahelised esinemissagedused, mis viitab populatsiooni struktuuri erinevusele isegi eel juhul, olulisele võrreldavail tüüpidel jätta võrdlusel kõrvale üksikisendite esinemiesagedus. Üheteistkümne aasta jooksul on seda jaotumuetüüp! esinenud ainult kahel aastal ja ainult Ida-Eesti rajoonides, mis viitab sellise jaotumustüübi kujunemise võimalikkusele põtrade massilise immigratsioon! korral. Antud jaotumustüübi esinemise usaldatavust kui suurendab selle tüübi esinemine aastal kahes naaberrajoonis, kuhu on võimalik idapoolne immigratsioon - jaotumuste kokkulangevuse usaldatavus on suur = 1,90, df = 4; P> aastal vaadeldav jaotumuetüüp esines ainult Alutagusel (D rajoon), kusjuures kokkulangevus aasta jaotumusega on väga suur (X 2 = 1,54, df = 4» P > 80 %) aastal on D rajoonile kõige sarnasem jaotumus samuti naaberrajoonle E ja viimase naaberrajoonis С - s. o. LÕuna- Eesti (üksikute esinemissagedus on vastavalt 37 % ja 33 %, samal ajal kui Lääne- ja PÕhja-Eesti rajoonides see oli 26% ja 27 %). Kasutades aditiivsuse reeglit (et arvestada C, D ja E rajoonide jaotumuste vaheliste seoste usaldatavust), ilmneb nende kolme rajooni väiksem usaldatavus vahelise seose märksa = 18,03, df = 12; P >10 %) võrreldes В, С ja E rajoonide vaheliste jaotumuste seostega (X 2 = 9,49, df = 12; P> 60 %), mis ei võimaldagi I jaotumuetüüp! lugeda levinuks peale D rajooni ka E ja С rajoonis, kuigi mingi antud jaotumustüübi "mõju" neie rajoonides ilmneb. I I j a o t u m u e t ü ü p o n v a a d e l d a v a l a j a v a hemikul kõige sagedamini esinenud ja allee aastast alates on see täielikult asendunud teiete tüüpidega. Arvesta-

, samal ajal aga Lääne-Bestis ja Vahe-Besti põhjaosas")
45 dee seile jaotumustüüpi pikaealieuat, võib seda pidada "normaalseks" väljakujunenud territoriaalsete ja sotsiaalsete suhetega põdrapopulatsloonile iseloomulikuks Isendite jaotumuseks. Vaatlusperioodi esimesel neljal aastal oli vaadeldavast jaotumustüübist oluliselt erinev ainult Lääne-Bestis aastal täheldatud jaotumus aastal hõlmab II jaotumuetüüp Vahe- ja Lääne-Besti, kusjuures antud rajoonide (A, B, C) jaotumuste Ohtlikkus on suur (summaarne 3^2-9.04, df * 15; P > 80 %) aastal taastub Ida-Bestie II jaotumuetüüp (rajoonide C, D, E jaotumised on usaldatavalt kokkulangevad - summaarne if «2,56, df = 12; P>99,5 %), samal ajal aga Lääne-Bestis ja Vahe-Besti põhjaosas ilmneb suuremate karjade osatähtsuse tunduv tõus ja vastavalt V jaotumustüübi kujunemine aastal ilmub Alutagusel uuesti erakordselt rohkelt üksikuid (I jaotumuetüüp), kuld Vahe-Besti põhjaosas taastub II jaotumuetüüp; rajoonide В, С ja В jaotumuste Ohtlikkus on võrdlemisi suur (summaarne X 2-9,49, df 12; P > >60 %) aasta eri rajoonide jaotumuste ühtlikkue asus veel usaldatavuse piires ning detailanalüüeil ilmnes, et ainult Lääne-Besti jaotumus erines teiste rajoonide omast tunduvalt - selle tõttu ka II ja talle väga lähedase III jaotumustüübi vaheline piir pole sel aastal kuigi terav aastal on usaldatavalt mitteerinevate jaotumustega Lääne- ja Vahe-Beetl (summaarne xf - 8,94, df 18; P > 95 St) ning nende rajoonide Jaotumus surn randmete alusel on Ilja III Jaotumustüübi vahepealne, Ida-Beatl (rajoonid D ja B) jaotuauekõver on aga II ja I jaotumustüübi vahepealne aastal säilib II jaotumuetüüp veel ainult Xagu-Bestis. III jaotumuetüüp on võrdlemisi lähedane II tüübile, kuld võrreldes nende JaotumuetttOplde aii nurandmeid, on erinevus siiski kõrgeimal taeemel usaldatav (X^2 * = 36,37***, df - 8). See muidugi ei välista nendele Jaotumuetüüpldele vahepealee jaotumuse esinemise võimalust, nagu ongi Ja aastal aastal laieneb vaadeldav Jaotumuetüüp ka Alutaguse rajooni ning Lääne-Beetle kujuneb jaotumuetüüp; III Jaotumustüübiga В, С ja D rajoonide jaotumus pole väga ühtlik (summaarne = 20,22, df - 21; P>50$>). I V J a o t u m u e t ü ü b i l e o n i s e l o o m u l i k e r a kordselt väike üksikute esinemissagedus. See tüüp on eelne 44 V
. 1971. aastal seevastu JaotumuskÕverate väga tugev kokkulangemine kahes naaberrajoonis (X?")

46 nud ainult kahel aastal kolmee rajoonle. On tõenäoline, et 1963«aasta Lääne-Besti andmete kokkulangevus aaeta andmetega on juhuslik, kuna 1963» aastal oli analüüsitavate karjade arv suhteliselt välke (62 karja) aastal seevastu JaotumuskÕverate väga tugev kokkulangemine kahes naaberrajoonis (X? = 2,58, df - 7; P>90 %) ei eaa olla juhuslik ning Õigustab antud jaotumustüübi püstitamist. V j a o t u m u s t t i ü b i l e o n i s e l o o m u l i k suuremate gruppide tunduvalt suurem osatähtsus kõigist taietest tüüpidest aastal esines Lääne- Ja PÕhJa-Bestls - kahes naaberrajoonis. Jaotumus on ieendiliete gruppide osas väga lähedane II jaotumustüübile, kuld suuremate gruppide osas kokkulangev V tüübiga, mille alusel need rajoonid ongi paigutatud sellesse tüüpi aastal säilib sama jaotumus ainult Lääne-Bestis, kuigi PÕhJa-Besti Jääb jaotumuse iseloomult kõige lähedasemaks aastal püsib V jaotumuetüüp Lääne-Bestis, kus 1969 aastal asendub III Jaotumustüübiga, mis ei hõlma aga mitte ainult L*tne- vaid ka Vahe-Bestlt aastal on Lääne-Bestis Jällegi V jaotumuetüüp ja Lääne-Besti on Jällegi samas erandlikus, teistest rajoonidest erineva jaotumusega seisundis nagu ta oli ka ja eriti aastal aastal asendab Lääne-Bestis V tüüpi IV-ga ja V jaotumuetüüp hõlmab kogu Ida- ja LÕuna-Eestl, kuld jaotumus on neis rajoonides võrdlemisi ebaühtliki summaarne - 32,10 (df 24) ning Jaotumuste ühte kollektiivi kuulumise usaldatavus on 20 Ja 10 % vahel. Kuna LÕuna-Besti (C Ja B) rajoonide jaotumuste vaheline seos on märksa tugevam kui zlende rajoonide Jaotumuste seosed Alutagusega, võib LÕuna-Besti rajoone vaadelda koos - sel Juhul nende rajoonide jaotumuse erinevus Alutagusel täheldatavast on usaldataval tasemel. LÕuna-Besti rajoonide jaotumuste lähem võrdlus lubab lugeda neid IV Ja V Jaotumustüübi vahepealeelka, Alutagusel esinenud jaotumus on aga V Ja III Jaotumustüübi vahepealne Jaotunmstüflpide.1 a nende esinemise analüüs PÕdrapopulatsiooni arvukuse Ja asustustiheduse pideva suurenemise tingimustee, nagu see oli vaadeldaval ajavahemikul, võib oletada, et sellele kaasneb suuremate gruppide 45


47 osatähtsuse suurenemine - mõned autorid /175, 200/ nimetavad gruppide suuruse olevat otseses seoses populatsiooni arvukusega. Teiselt poolt muutub asustustiheduse suurenedes järjest tõenäolisemaks ühes ajus mitme grupi esinemine, mis metoodikaveana samuti suurendaks suuremate gruppide osa. Siinkohal lähemalt analüüsimata, kummast põhjusest tingituna, kuid vaadeldaval ajavahemikul võime (välja arvates vähesed erandid) konstateerida vaatlusperioodi teisel poolel selliste Jaotumustütipide levikut, milles suuremate gruppide osatähtsus on suurem kui vaatlusperioodi esimesel poolel esinenud Jaotumustüüpides. Viie püstitatud jaotumustüübi järjestamisel lähtutigi mitte nende ajalise esinemise vaid loogilise arengu järjekorrast. Arvestades kõiki jaotumustütipe võime nentida, et eri suurusega gruppide esinemissagedus võib kõikuda suurtes piirides (Joon. 39): üksikute osa , üksikute ja kaheliste osa kokku - 37 %, üksikute, kaheliste ja kolmeliste osa kokku , mis võib moodustada kõigist karjadest ! Kui sellise diapasooniga jaotumised esineksid läbisegi eri aastatel ja eri rajoonides, võiks põdra karjasuet pidada mür läkski väga labiilseks, determineerimatuke või väga paljudest muutlikest teguritest mõjutatavaks nähtuseks, millel pole olulist tähtsust populatsiooni elus. Kuna aga kogu muutlikkuse diapasooni hõlmavad eri Jaotumustüübid esinevad küllaltki seaduspäraselt üheaegselt mitmetes rajoonides või ka kogu territooriumil ja säilitavad järjepidevuse real aastatel, siis on meil põhjust pidada neid jaotumustütipe reaalselt kehtivaiks ning, analüüsides neid lähemalt, püüda selgitada nende tüüpide erinevuste olemus ja erinevuste põhjused. Alustagem äärmuslikest j aotumustüüpide st, mille esinemissagedus Ja ka levik on olnud kõige tagasihoidlikumad. I ja II jaotumustüüpi lähemalt võrreldes selgub, et nende põhiline lahkuminek seisneb üksikute ja kaheisendiliete gruppide esinemissageduste olulises erinevuses, kusjuures teiste gruppide omavahelised suhted on enam-vähem kokkulangevad (joon. 41) ning ei luba neid jaotumus! lugeda usaldatavalt erinevaiks (5^2 = 4,89, df = 4; P > 30 %). Järelikult võime I ja II jaotumustüüpi lugeda põhiliselt sama struktuuriga tüüpideks, kuid nende erinevuse on põhjustanud li'


48 suure arvu Ukalkute.1a kahelaendillate gruppide juurdetulek USM. IV ja V jaotumuetuubl oluline erinevus seisneb ainult üksikute osatähtsuses: arvates need võrdlemisel välja, ilmneb kõigi teiste gruppide arvuliste suhete väga tugev kokkulangemine (%?» 4,20, df = 7; P > 70 56), mis ei luba neid mingil juhul pidada pärinevaike eri kollektiividest. Järelikult on IV jaotumuetüüp kujunenud V tüübist suure osa üksikute lahkumise tagajärjel. % I г в 9 is/gr. Joon Eri suurusega põdrakarjade esinemisaagedused oktoobrikuu jaotumustüüpidea, kui üksikisendid on välja arvatud. II ja III jaotumustüübi vaheline erinevus seisneb viimases nelja ja enama isendite arvuga gruppide suuremas osatähtsuses, kuna isendiliste gruppide osa on peaaegu täielikult kokkulangev (joon. 39). 47

 esitamisega, mis aga võib olla täpselt sama täiesti erinevate jaotumuste korral.")
49 Arvestades eelnenut, võib põhilisteks jaotumustttüpldeks pidada teist ja viiendat, kusjuures kolmas tüüp on üleminevaks teiselt viiendale Ja esimene ning neljas tüüp on kujunenud populate!oonisiseste liikumiste tulemusena JAOTOMBST MÜHAVAD tkovbxd PÕdrakarjade jaotumuse kohta areaali eri osades on suhteliselt vähe andmeid, mida oleks võimalik kasutada võrdlemiseks: paljudel Juhtudel esitatakse eri kuude ja rea aastate andmed liidetuna või piirdutakse ainult karja keskmise suuruse (e. karjasuse koefitsiendi e. karjasuse näitarvu) esitamisega, mis aga võib olla täpselt sama täiesti erinevate jaotumuste korral. Häiteks Tambov! oblastis /211/ on 12 aasta jooksul märtsikuudel kogutud ulatuslikud andmed (andmed 6964 karja suuruse kohta), mis on aga esitatud koondina. Võrreldes neid andmeid Eesti detsembrikuu andmetega, Ilmneb erinevus nii a. kui ka a. jaotumustes: vaatlusperioodi algaastate ja ka viimase aasta detsembrikuu jaotumuste erinevus Tambovl jaotumusest on usaldatav kõrgeimal tasemel (3^2 «42,92***, df «8 ja 3$^ = 44,60***, df=8). Mõlema võrdluse puhul on kõige suuremad erinevused üksikisendite esinemissageduses, mis on oluliselt suurem Eesti andmetes. Jättes need jaotumustest välja, väheneb tunduvalt jaotumuste erinevuste olulisus (9^ vastavalt 15,44* ja 20,51**, df=7). Tambov! jaotumuses on märgatavalt rohkem 2- ja 3-isendi li ei gruppe, isendiliste gruppide esinemissageduses on erinevused kõige väiksemad ning a. Eesti jaotumusega võrreldes on nende gruppide eeinem!esagedused täiesti kokkulangevad. Sende areaali eri osade jaotumuste oluline erinevus on kõige tõenäolisemalt tingitud andmete pärinemisest eri kuudelt - sellele viitab eriti üksikute tunduvalt väiksem osa Tambovl Jaotumuses, teiselt poolt võis erinevust suurendada ka erineva asustustiheduse Ja erinevate tingimustega aastate materjalide liitmine. Eesti naaberaialt Leningradi oblastist on kirjanduses toodud andmed põdrakarjade jaotumusest aastatel summeerituna kuude lõikes /190/. Nagu näitas Eestist samalt 4ö
.")

50 ajavahemikult pärinevate oktoobrikuu jaotumuste võrdlus, ei erinenud need omavahel oluliselt, mie lubab neid eamuti kooe vaadelda. Leningradi oblasti ja Kesti oktoobrikuu jaotumuste erinevus ei küüni olulisele tasemele, kuid ei erine sellest ka palju (^2 = 10,79, df = 6; P>5 %). Põhiline lahkuminek seisneb Leningradi oblastis viie- ja enamaleendlliete gruppide väiksemas osatähtsuses võrreldes samadel aastatel Bestie täheldatugai vaadeldes suuremaid gruppe koos, suureneb jaotumuste sarnasus tunduvalt (^2 «3,09, df- 4;P>50jQ. Võrreldes Leningradi oblasti andmetega samade aastate Alutaguse andmeid, ilmneb neis kokkulangevus ka suuremate gruppide esinemissageduses ning jaotumus ei erine eduliselt (X ,66, df - 5; P>40 JÜ lesti detsembrikuu jaotumuse et võs> koos vaadelda a. andmeid, millede erinevus Leningradi oblasti jaotumusest on tõenäoline juba 99,5 % tasemel (з; 2-20,69**, df - 7) ning tingitud Jällegi suuremate gruppide suuremast osatähtsusest Bestiet vaadeldes viie ja suurema isendite arvuga gruppe koos, vsib Jaot usual lugeda kokkulangevaiks (Ц 2» 3.99, df - 4; P > 40 %). Võrreldes Leningradi oblasti jaotumusega ainult Alutaguse andmeid, ilmneb nende väga suur sarnasus (^2-3,32, df - 5; P>60<). Järelikult aastatel oli Leningradi oblastil» Iseloomulik põdra karjade.jaotumus levinud k«basti Idaossa Alutagusele. mis külgneb Leningradi oblastiga. Alutaguselt lääne poole tõusis jaotumuses märgatavalt suuremate karjade osatähtsus ja vähenes üksikute osa. Eelnenust võib teha veel teisegi järeldusel sama kuu.1a aasta laotumused võivad olla kokkulangevad nii suurtel aladel kui ka teatud t<n^mmt«i erineda isegi sellise väikese terrltoor»' ^, "«ro seda Eesti, piires. Teguritest, mis tingivad põdrakarjade jaotumuste erinevusi, käsitlesime Juba klimaatiliste tingimuste mõju (tõenäoliselt koosmõjus toltumistingiaugte muutumisega) karjasuse suurenemisele oktoobrist detsembrini, mis on kirjanduse andmeil kestab edasi kuni talve lõpuni /32/. Samal ajavahemikul, samade klimaatiliste tingimuste korral eri territooriumidel (või samal territooriumil eri aastatel) jaotumust määravatest teguritest analüüsiksime alljärgnevalt (1) hüpoteesi, et Isendite arv gruppides on juhuslik ning gruppide jaotumus on selle tõttu vaetav juhuslikule 7 49 (Poiesonl)
 - Õigemini valem, mis võimaldab arvutada eri suurusega gruppide esinemise")
 vastavad Polasonl jaotusele.")
51 jaotumusele; (2) arvukuee Ja asustustiheduse mõju jaotumusele; (3) areaalleleeete llllcumlete ja(4) populateloonl ootelaalee struktuuri mõju jaotumusele. K.D. ZSkovi Ja J.B. Sapetini /127/ poolt eeltatl põdrapopulatsiooni jaotumuse matemaatiline mudel (nagu väljendatud artikli pealkirjas) - Õigemini valem, mis võimaldab arvutada eri suurusega gruppide esinemise teoreetilise sageduse. Teoreetilise Jaotuse võrdlemine tegelikuga võimaldake hinnata loendusandmete Ja nende andmete ekstrapoleerimise Õigsust. Valemi tuletamisel lähtuti eeldusest, et nii põdragrupplde esinemine loendusalal kui ka isendite arv «trupis (eri suurusega gruppide esinemissagedus) vastavad Polasonl jaotusele. Sellele seisukohale tugines ka Y.S. Smirnov ulukite loenduste matemaatilise analüüsi käsitlusel /194/ ning J.A. Qerassimovi /111/ andmete kriitilisel vaatlusel /193/. Kontrollimisel Eesti materjalil selgus, et ühelgi jaotumustüubll ühelgi kuul polnud tegelik jaotumus kokkulangev Poissonl Jaotusega. Et Jaotumuet võivad oluliselt mbjutada nn. nullandmed, s. t. nende proovialade arv, millel p8tru ei esinenud, arvutati tegeliku jaotumuse jaoks ka "nullide" arv, mis peaks seal olema, kui tegu oleks Poissonl jaotusega*. Kuld Isegi sel juhul tegeliku jaotumuse erinevus Poissonl Jaotusest Jäi kõrgeimal tasemel usaldatavaks. Järelikult sügis-talvisel perioodil Isendite arv gruppides pole juhuslik, nad pole levinud ühtlaselt ja üksteisest sõltumatult ning sel põhjusel ka nende jaotumus ei vasta Poissonl Jaotusele. PSdra karjade suuruse seose kohta populatsiooni arvukuse Ja asustustihedusega on avaldatud täiesti vastandlikke seisukohti: "PStrade karjasuse koefitsent (näitarv) on põtrade asustustihedusega võrdeline. Seal, kus loomade arv on suur, kohatakse sagedamini suurema isendite arvuga gruppe, mis on omakorda seotud nende koondumisega toitumlsaladele." /200/. Aeuetuetlheduse määravat msju karjade x Poissonl Jaotust iseloomustab keskmise (M) ja ruuthälbe (s 2 ) väärtuste kokkulangemine, järelikult 50
 Ja suurte p8dragrappide arv ei ole seoses nende üldise arvukusega.\" ^11/.")

52 suurusele rõhutatakse ka Leningradi oblastis /175/ ning otse vastupidist väidetakse Tambovi oblasti materjalide analüüsil : "... karjasuse koefitsient (näitarv) Ja suurte p8dragrappide arv ei ole seoses nende üldise arvukusega." ^11/. Eestist pärinevate oktoobrikuu and*ete analüüsil vbib sedastada, et abundantsi Ja tiheduse korrelatsioon on positiivne ja kõverjooneline: kui tihedus on allpool nn. piirväärtust, pole abundantsi muutused alati tiheduse muutustega samasuunalised Ja alles siis, kui tihedus ületab piirväärtuse, suureneb Järsult samasuunaliselt ka abundants. See "piirväärtus" on ühtlasi frekvents! Ja tiheduse väga tugeva positiivse sirgjoonelise korrelatsiooni murdekohaks, mille Järel frekvents! juurdekasv Järsult aeglustub, võrreldes tiheduse juurdekasvuga. Siit võime teha Järelduse, et tiheduse "piirväärtus" tähistab sellise asustustiheduse saavutamist, mille puhul on asustatud suurem osa elutlnglmustelt sobivaid alasid ja ühte ajju hakkab jääma sageli rohkem kui üks grupp. See vastab uue jaotumustüübl kujunemisele, milles suuremate gruppide osatähtsus (ja Järelikult ka abundants) on suuremad kui oli väiksema asustustiheduse puhul. Seega meie andmetel sõltub abudantbi (karjasuae koefitsiendi) ja tiheduse seose tugevus populatsiooni tiheduse tasemest ning Õiged on nil B.N. Teplova kui ka V.D. Heruvlmovl seisukohad: Okaa looduskaitsealal, kus andmeid kogus В Л. Терlova, on põtrade asustustihedus ilmselt tunduvalt "piirväärtust" ületav ja märksa kõrgem kui Tambovi oblastis, kust pärinevad V.D. Heruvlmovl materjalid. Põdra suur areaalieieene liikuvus on üldiselt teada ja seda on täheldatud nii möödunud kui ka käesoleval sajandil korduvalt ka meie aladel. Kida tihedam on põtrade levik mingil territooriumil ja mida suurem on nende arv, seda raskem on märgata põtrade populatsioonisiseseid liikumisi parasvöötme aladel, kus ei esine massilisi etigis-talviseid ja kevad-talvissid rändeid. Teatud pidepunkte populatsioonisiseete liikumiste selgitamiseks ka suhteliselt suure asustustihedusega aladel annavad vaatlused põdra areaali servaaladslt või piirkondadest, kus põdrad on levinud sporaadiliselt, ning Isegi vaatlused nende ilmumisest linnadesse. Hii näiteks S.A. Hovikovi Ja P.D. Ivanovi /176/ andmeil registreeriti aastatel põdra ilmumine Leningradi. 51
 oli nende hulgaa 73,7 * ning valdav enaaue (87,4 %) linnapiiridesse Haualei tolmus mais-juuniв - põdralehmade poegimise ajal, millal nad ajavad eemale endi eelmisel aastal sündinud")

53 Enamik nelet olid üksikud, harva leendllleed ja veelgi harvem leendllleed grupid. Hoori (aastasi) oli nende hulgaa 73,7 * ning valdav enaaue (87,4 %) linnapiiridesse Haualei tolmus mais-juuniв - põdralehmade poegimise ajal, millal nad ajavad eemale endi eelmisel aastal sündinud vasikad. Ka Moskva piiridesse põtrade ilmumise juhtudest langeb 68 % maile-juunile, augustis ilmusid vaid vähesed ja uuesti suurenes immigrantide arv septembris /122/. Lisamata analoogseid andmeid põdra kohta, vblb veel märkida, et pbder ei kujuta endast mingit erandit teiste imetajatega võrreldesl reeglina on kõige liikuvamad noorloomad "iseseisva elu" alustamisel ning noorloomadest on vähim еша- 1оomaga seotud isased /216/. Jaotumuses peaksid massilisemad populatsioonisisesed noorloomade liikumised kajastuma üksikute osatähtsuse suurenemises immigratsloonialadel Ja üksikute osatähtsuse vähenemises emigratsloonialadel. See saab aga märgatav olla ainult sel juhul, kui emigratsiooni- ja lmnigrataioonlalad on erinevad territooriumid i enam-vähem sarnase asustustiheduse Ja sama sigimise edukuse korral peaks vastasel Juhul kbikldel aladel immigrantide arv vbrduma emigrantide arvuga ning mingeid märgatavaid muutusi Jaotumuses ei esineks. Viimasel juhul oleks loogiline oletada teatud kbrvalekaldumlsi Jaotumuses ainult füüsiliste barjääride ees. Vaadeldaval ajavahemikul vblb Eestis immigratsioonist tingituna Jaotumuse kbrvalekallet täheldada 1965» ja aasta oktoobrikuudel Ida-Bestis, millal seal esines nn. I jaotumustüüp (vt. lk. 43). Teistel aastatel pole immlgrateloon olnud nii arvukas vbi sellise üksikute enamusega, mis oleks suutnud oluliselt mbjutada kohapealee populatsiooni "normaalset" Jaotumust. Lääne-Eeetis peaaegu pidevalt täheldatud teistest aladest erinevaid jaotumuei vblb enamusel aastatel pidada tingituks "normaalsest" populatsioonisieeseet liikuvusest, aastal aga eelmisel aastal Ida-Eestisse jõudnud "levikulaine" Jõudmise st Lääne-Eestisse. Kuna siin on füüsiliseks barjääriks meri nii pbhja- kui ka läänesuunas, on siin kõige tõenäolisem olulise osa migrantide koondumine. Sellest tingituna olulised erinevused karjade jaotumuses ja ka teistest aladest varasem suurema asustustiheduse ilmumine. Populatsiooni sotsiaalse organisatsiooni mõju selgita 52
.")
, siis on loomulik, et karjade Jaotumuses on oluline osa vasikate arvul ühel emaloomal ja vasikate osatähtsusel populatsioonis üldse.")
54 miseks arvutati korrelateioonid rea sotsiaalselt erineva tähtsusega grupeeringute arrukuee hing mõningate eiglmlsnäitajate Ja erineva suurusega pftdrakarjade esinemlssageduse vahel. Kasutati ainult «aastate andmeid, millal frekvents polnud J8udnud veel "piirväärtuseni*, millest aiatee hakkavad ilmnema "normaalset" Jaotumist rikkuvad nähud (tabel 51). Kuna p8d raitarjade tuumikuks on reeglina nn. ema perekond (vt. lk. 11), siis on loomulik, et karjade Jaotumuses on oluline osa vasikate arvul ühel emaloomal ja vasikate osatähtsusel populatsioonis üldse. See ilmneb ka olulistes korrelatsioonides vasikate esinemise protsendi Ja karjade suuruse vahel: mida rohkem on populatsioonis vasikaid, seda väiksem on üksikute esinemissagedus oktoobris Ja nommbrls ning seda sunrem on suuremate karjade osa detsembris. Hovembrls kui Ulemlnekukuul on vasikate eslnemlesagedus positiivses korrelatsioonis 4,..5-lsendlllste Ja negatiivses korrelatsioonis 6- Ja enamalsendiliste gruppide esinemissagedusega. Analoogsed on seosed jaotumusega kahe vasikaga emaloomade esinemissagedustel, vasikata emasloomade osatähtsusega on aga Jaotumusel seosed vastupidised, nagu on ka loogiline oodata. 53


55 6. POPULATSIOON BIOLOOGILISE SÜSTEEMINA JA TA RATSIONAALNE KAS VT AMINE 6.1. POPÜLATSIOOH JA TA АШЕПГОТВ piihituarare Seoses populatsioonide uurimise tulipunkti tsuseisega - selle üheks iseloomustuseks on ka ökoloogia nimetamine teaduseks populatsioonidest /214/ - suurenes järsult zooloogilist e tööde arv, milledes käsitletakse nii populatsiooni kui ka sellest ülemaid Ja alamaid at rukt uuri tihi kuid. Paljudel Juhtudel ei määratleta täpsemalt kasutatavaid termineid,teistel Juhtudel aga samale terminile antav sisu v8ib tunduvalt erineda /174, 187, 217/. Asudes Eestis kogutud Ja eespool esitatud konkreetse materjali lahtimõtestamisele populatsiooni struktuuri seisukohalt, on vajalik eelnevalt peatuda populatsiooni struktuuriühikute käsitlemisel (eriti euurimetajate materjalil) püüda selgitada erinevaid struktuuriühikuid ning iseloomustavad pshllised tunnused ja samaväärsete ühikute eristamise printsiibid. Populatsiooni iseloomustava tunnusena arvukuse dünaamika rõhutamine esineb mitmel autoril. S.S. Schwarz /215/ kirjutab! "... populatsiooniks ranges msttes tuleb nimetada mikropopulatsloonide kogumit, mis on omavahel seotud tihi ee päritoluga (reservaatidest) Ja arvukuse нииджпи*«шя1;е seaduspärasustega" (allakriipsutus minu, H.L.). Ja edasis "On näidatud, et mitte ainult sigimise fonoloogia erinevus (ammutuntud fakt), vaid ka erinev arvukuse dünaamika tüüp v8ib naaberpopulateioonld kindlalt isoleerida". Samaaegselt mööndakse ka vsimalust, et arvukuse dünaamika vsib olla erinev mikropopulatsioonides ja naaberpopulat ei oonide e aastal väljendab sama autor oma seisukohta populatsioonide piiritlemise küsimuses järgmiselti "Populatsiooni piiride selgitamine on seotud suurte raskustega, mis tuleneb pshlliste populatsiooni protsesside keerukusest ja dünaamilisusest. Käesoleval ajal aaaafc p^mald tulemusi populatsiooni piiride üraaiaa ддцдвма düaaaaüjta цщнода. aübüal, mida 54

.")
56 täiendavad geneetilised Ja morfoloogilised uurimused/122/ (allakriipsutus sinu, H.L.). B.A. Larini /1*5/ eelaukoht on lähedane: "Rakendusökoloogias, kus uurimise põhieesmärk on arvukuse dünaamika uurimine, ta muutuste prognoosimise rõlmaluste selgitamine Ja lõpptulemusena arvukuse dünaamika Juhtimine^ mõistetakse populatsiooni all tavaliselt sellist ühe liigi isendite elementaarset grupeeringut, mille lndivlldldevahslised vastastikused Beosed võimaldavad säilitada üht või teist arvukuse taset." (allakriipsutus minu, H.L.). 18eed aastad hiljem autor täpsustab: "Praktilises plaanis tuleb populatsiooni kui inimese poolt m8jutetava elementaarse objekti all mõista sellist gruppi ühe liigi isenditest, kelle interaktsioon võimaldab taastada arvukuse flhe-kahe reproduktiivse seseonl vältel. Sellise grupeeringu piiride määramisel on esimeseks kriteeriumiks aktiivsuse raadius - isendite ümberasetumlse keskmine kaugus eriti sigimiseks valmistumise perioodil." /146/ (allakriipsutus minu, H.L.). S.S. Schwarz /216/ toob populatsioonide ja nende pilritlemiee ühe näitena nende laboratooriumi poolt Taga-üraall metsast epi järvedel (vahega 0, km) tehtud uurimuse ondatrast. Selgus, et isegi sarnastel järvedel arvukuse dünaamika oli erinev, kusjuures arvukuse dünaamika määras põhiliselt erinevus noorte ellujäämises] "Esitatud andmed näitavad, et vahetus läheduses asuvate Järvede ondatra asurkoidadel on erinevad arvukuse dünaamika seaduspärasused ja neid v8ib vaadelda kui mlkropopulatsioone..." (lk. 97). z Arvukuse dünaamikale asetavad lõpptulemusena populatsioonide eraldamisel rshu ka Olmelfarb jt. /112, lk. 13/1 "Himelt isendite taastootmise seadused, nende sündimine, ellujäämine v8i suremine on nendeks põhilisteks "jõududeks", mis viivad populatsiooni struktuuri ümberkorraldustele ta arvukuse muutustele. Sellepärast, uurides sellelaadseid protsesse, peame me termini "populatsioon" all т81р*и"» bin. loogiliste Indiviidide kogumit, kes asustavad teatud territnnf-iiinh vjh»wntnnrhinii ja kes on seotud taastootmise Ja ellujäämise protsesside ühtsusega." (allakriipsutus minu, ja x Autori kogemused /2/ võimaldavad ondatra arvukuse dünaamika erinevuste pshjusena lähestikku asuvates veekogudes näha eelkõige kiskjate ja talvetinglmuste mõju, mis lühiajalistel uurimustel Taga-öraalis jäid märkamata. On mõistetav. et nende tegurite mõju korral mingeist "mikropopulateloonidest" kõnelda pole võimalik.

, et \".")
57 H.L.). Samad autorid v8tavad populatsioonide kohastumise kvantitatiivseks hindamiseks kasutusele nende natallteedl - keskmise järglaste arvu tihe vanalooma kohta ajaühikus. Eelnenuga kokkulangev on I.V. Timofeeff-Hessovsky Jt./203, lk. 121/ eelaukoht: "... rieturni ee süsteem Ja (ttldleeaal kujul) sigimise edeteem on üheks tähtsamaist populatsiooni kohastumist iselooaustavaist tunnustest ning v8ib-olla selle muutlikkus eri populatsioonides väljendab liigilist evolutsioonilist plastilisust." Asudes käsitlema populatsiooni dünaamikat ruumis Ja ajas, tunni et arad samad autorid (lk. 43), et "... isegi ksige laiemalt levinud ja Üldtuntud liikidel pole selgitatud populatsioonide piire suhteliselt väikestel ja ülevaatlikel territooriumidel," ning edasi: "... ainult vähestel Juhtudel on uuritud üksikute populate!oomide piire, peaaegu mitte midagi pele teada naaberpopulataioonlde Tähelisest isolatsiooni astmest ja populatsloonidevmhelisest vahetusest looduslikes tingimustes ning samuti populatbioonills-dflnaetil 11 ste protsesside kvantitatiivsest küljest." (lk. 84). Selle peaaegu täieliku uurimatuee tingimustee en autorid siiski teinud üldistusi ja toonud näiteid populat ei oonide st, millest selgub nende arusaam nii populateioonlde suurusest kui ka nende piiride kohta. Imetajate populatsioonidest märgitakse mügri (Arvicola ka terrestrip) kohta, et üksikud populatsioonid h8lmavad territooriumi ühest kuni m8nekümne hektarini (lk. 46, 47) ning et paljude euurte imetajate populateioonid vbivad koosneda väga väikesest isendite arvust (lk. 47). Lindudest nimetatakse "headeks populatsioonideks" euurte kajakate üksikuid pesitsuskolooniaid Lääna-Siberl ja Kasahstani Järvedel,milles täi ska avanud isendite arv v81b ksikuda mitme kümne et mitmesaja paarini (lk. 47). "Populatsiooni hästi tuntud näiteks v8ib.nähtavasti pidada ka künnivarese (Corvue frugilegue) pesitsuskolooniaid" (lk. 47). Dendrolimus pinl populatsioonideks vsivat aga lugeda igal männil talvitunud isendite kogumit (mändide vahemaa 4 ml), nagu selgub lk. 28 toodud jooniselt. Populatsiooni piiride kohta väidetakse, et need vbivat olla nii teravad kui ka hajusad. "Teoreetiliselt oo populatsioonid piiratud teatud isolatsiooni pressiga; kuid praktiliselt isolatsiooni piiri pole mitte alati kerge selgitada Ja sellepärast tuleb juhinduda naaberpopulatslooni- 56


58 de asustustiheduse muutustest... asurkonna tihedus v8ib ühelt tipult tipule ninna üle madaldanud asustustiheduse oru*..." (lk. 24). Ja teisalt "K8ik need iga liigi asurkonna tiheduse tsentrid ongi reeglina populatsioonid " (lk. 36), sellele lisatakse siiski, et rahel nimetatakse neid ka 4ee*ldeke (1), kui neil pole iseseisrat eroluteioonilist saatust (?) ja kui nad on raid suurema ühiku osad (lk. 37). Tuginedes toitumise Ja slglmisnäitajate erinerusele, eristab V.D. Herurimor /211/ Tambori oblasti kola p&drapopulatsiooni. x J.P. Jasen /236/ nimetab, et "... Petftora ja Tjatka loomade /pstrade/ näitajad /nataliteet/ erinevad etatistiliselt usaldataralt, mis körruti kolju proportsioonidega on "Mr- nende populatsioonil! ee et Iseseisvu sect psdra euroopa alamlilgis," (allakriipsutue minu, H.L.). Analöüeides kelme eri laiuskraadi oblasti «nrtmnlri psdra»ekundäärsest mataliteedist (embrüone emalooma kohta), leiab ka I.D. Perovski /179/, et lõunapoolsete populatsioonide potentsiaalne riljakus on suurem ja stabiilsema iseloomuga kui põhjapoolsetel populatsioonidel. P.B. Jürgenson /231, 232/ on seisukohal, et "... tegelikult eksisteerir populatsioon peab asustama kindlat, küllalt püairatee piirides maa-ala, kuigi selliseid piire looduses selgitada pole kerge." /232, lk. 111/. Populatsiooni käitumine ja suhted keskkonnaga, rastusreaktsioonid, on ühetaolised. Vanuseline ja sooline struktuur võivad naaberpopulat ei oonidel kas erineda või ka mitte - selle tõttu pole nende erineruste alusel populatsioonide eraldamine Õigustatud. Vastandina S.S. Sohwarsi koolkonnale, kes raatarad lüki kui "... teineteisega samaräärsete ja praktiliselt sõltumatute populatsioonide kogumikku, mis on rsineliaed iseseisvalt eksisteerima vastastikuste mõjutusteta" /173, lk. 832/, on N.P. Haumor ja rida teisi ökolooge seisukohal, et liik on "... keerukas erineva tasemega hierarhiliselt üksteisele allurate populatsioonide kogum, kes on omavahel seotud indiriidide rahetusega: pidera vahetusega madalama tasemega populatsioonide rahel, perioodilise r6i episoodili- P.B. Jürgenson /231, lk. 52 / nimetab neid hästi isoleeritud elementaarpopulatsioonideks Tambori lokaalpopulatsioonis. > 57 8


59 aega kõrgema tasemega populatsioonide vahel." (lk. 832). N.P. Naumov /168/ on seisukohal, et andmete puudumine populatsioonide piiridest pole mitte populatsioonide puuduliku uurimise tulemus, vaid et reaalselt selged ph-m populatsioonide vahel looduseb ei esine. Selle põhjus seisneb populatsiooni kui ökoloogilise süsteemi kvantitatiivses omapäras: tsentraalse regulatsiooniaparaadi puudumises ja komponentide integratsiooni madalas tasemes. Selle tagajärjel populatsioonide piirid pole mitte järsud, vald üleminekuvööndid. Võttes populatsioonide eristamise aluseks arvukuse dünaamika (arvukuse dünaamika sama-ajallsuae), me ei piir^t^a mitte populatsioone, vald arvukuse dünaamikat suunavate tegurite mõjualasid: looduslikud tingimused on primaarsed, arvukuse dünaamika sekundaarne. Asjata ei tõstnud P.B. Jürgenson /231, 232/ uuesti üles A.N. Formozovi poolt Juba aastal kasutatud mõistet "sarnase sobivusega ala" (" область сходственной благоприятности среды"), mille all mõistetakse ala, mille ulatuses toimuvad olulisemate(populatsiooni arvukust määravate) tingimuste ühesugused muutused. "Sarnase sobivusega alade piiride määramine (mitte ainult ühe, vaid tähtsamate faktorite summa alusel) võimaldab põhjendatult selgitada üksikute liikide suurte looduslike 1 okaalpopulat sl oonide piirid", /231, lk. 57/ X. P.B. Jürgenson annab sellele suurema tähtsuse, kui mis tahes morfoloogilisele tunnusele. Analüüsides suurtel aladel (tundra, meteatundra) rea näriliste populatsioonide erakordselt nõrka diferentseeritust, väidab ka S.S. Schwarz /215, lk. 1465): "Tuleme järeldusele, et naaberpopulatsloonide vastastikune mõju soodustab nende konsolideerumist ja kui terviku progressiivset evolutsiooni. On loomulik, et meie ei tea selle tsooni piire, mis ühendab populatsioone tervikuks. Võib oletada, et need piirid määratakse samaste elutingimustega." (allakriipsutus minu, H.L.). Sobivate elutingimustega ala esinemise pidevust loex Sama autori monograafilises töös /232, lk. 131/ lokaal populat slooni asemel kasutatakse väljendit "suur populatsioon". 58


60 taksegi populatsiooni terviklikkuse tagatiseks Ja sellise ala piire - populatsiooni piirideks: "Öeldu sünteesina soovitame ksikide imetajate populatsioonide kaardistamisel tsmmata piirid maastiku v8l füüsilie-geograafilise tõkke piire mööda, kui see läbib maastikku", soovitab V.E. Flint /209/. Selgem pole olukord ka populatsioonide alaühikute määrangule, eristamises ega nende nimetustes. Erinevast terminoloogilisest ja sisulisest segadusest populatsioonisiseste etruktuuriühikute käsitlemisel aimavad iseloomuliku pildi spetsiaalse nõupidamise* materjalid, millest esitan ainult suurimetajate populatsioone puudutavad näited. Selleks, et pääseda populatsiooni alaühiku nimetamisel "populatsioonist", teeb V.E. Flint /208/ ettepaneku populatsioonile kohe Järgnev alaühik nimetada meeruseks (merus - ld. osa), millele Järgneks N.P. Naumovi /167/ soovitatud alaühik partsell. Meerust kasutab populatsiooni alaühikuna E.V. Ivanter /130/. Ukraina Karpaatides eraldatakse arvukuse alusel hirvel ja metskitsel kolm geograafilise populatsiooni tüüpi /120/. PÕhja-Kaukaasias elutsevad halljänesed jaotatakse nende elupalgalise leviku alusel kolme bioloogilisse gruppi /156/. Metskitsede arvukuse ja eluala reljeefi struktuuri. (!) alusel eristatakse Taga-Baikali lääneosas 12 territoriaalset grupeeringut, mis pidavat olema "spetsiifiliste tunnustega" - asustustiheduse ja karjaausega /195/. Artiklis esitatud andmete statistiline analüüa näitab asustustiheduse ja karjasuse tugevat korrelatsiooni (r s =to,8(? x, N = 12), mille tõttu "spetsiifilistest tunnustest" jääb järele ainult arvukus. Moskvalähedasel alal analüüsiti põdra kohalikke populatsioone, edaspidi räägitakse ka Moskvalähedastest põdrapopulataioonideat /124/. Omapärane on populatsioonide käsitlus N.V.Rakovil/184, 185, 186/, kes uuris Ussuurimaa põdrapopulatsiooni. Autor Nõupidamine teemal "Liigi populatsiooni struktuur Imetajatel" (M., 1970), korraldajaike Moskva Looduauurijäte Selts, Moskva Ülikooli Bioloogia-Mullateaduskond Ja zooloogia kateeder. 59
 alusel ning «ristab üssuurimaal 31 põdra geograafi 11 at populatsiooni (geneetiliselt vähesegunevad) Ja 98 ökoloogilist populatsiooni (geneetiliselt segunevad).")

61 peab võimalikuks põdra Jakuutia Ja üeeuurl alamliigi erlstaulat viljakuse (IT) Ja sarvede kuju (!?) alusel ning «ristab üssuurimaal 31 põdra geograafi 11 at populatsiooni (geneetiliselt vähesegunevad) Ja 98 ökoloogilist populatsiooni (geneetiliselt segunevad). Teades, et p8dra viljakus Ja sarvede kuju on suuresti muutlikud, pole võimalik uskuda, et need võiksid olla suure arvu populatsioonide usaldava eristamise aluseks. Autori järeldusi toetava faktilise materjali ja selle analüüsi puudumine nimetatud töödes sunnib H.V. Bakovi seisukohti pidama spekulatiivseteks. P.3. Jürgensoni /231, 232/ seisukohad ja ta alajaotuste eristamiseks on Järgmised. populatsiooni Väikseimaks populatsiooni alajaotuseks on elementaaroopulatsloo», mida enam pole võimalik jaotada. Asünkroonsus arvukuse dünaamikas Ja teistes elulistes ilmingutes on üks eri elementaarpopulatsloonidesse kuulumise tunnustest Ja selle reaalsuse kinnitus. Mitu elementaarpopulatslooni võivad moodustada lokaalpopulatsiooni*. Blementaarpopulatsiooniks on tedrel ja metsisel nende mängukooslused (pindalaga ha), põdral võivat elementaarpopulatsioon hõivata teatud juhtudel mitusada tuhat hektarit ning aellee esineksid ainult perekondlikud või talvel ka etruktuuritud grupeeringud, arvukusel pole elementaalpopulatsioonlde piiritlemisel määravat tähtsust. Põdra elementaarpopulatslooni all tuleb mõista isendite gruppi, (1) kes asustavad sama ala, mille piiridest nad kogu aasta jooksul ei välju (välja arvatud liigsete noorloomade emigratsioon) Ja (2) kellel on ühine käitumine ja sesoonsed migratsioonid selle elementaarpopulatslooni areaali piirea. Tõenäoliselt taolise elementaarpopulatslooni näitena kirjeldabki sama autor /232, lk. 116, 117/ Moskva oblastis 200 ООО ООО ha pindalal eelnevaid põdra grupeeringuid, mida ta nimetab küll populatsioonideks. Populatsiooni jagunemist elementaarpopulataioonlke peab Õigeks ka J.P. Jazan /236, lk. 148/: "Elementaarpopulatsioon on põtrade grupeering, kee tundlikult ja ühemõtteliselt reageerib keskkonna muutustele üldse ja hääbumisele eriti." toidubaasi x Lokaalpopulatsiooni ei märgita monograafilises töös /232, lk /. 60


62 Vaadeldes pool toodud seisukohtade valguses Seati materjale psdra populatsiooni struktuuri ja ta produktiivsema dünaamikast, vbiae raita, et -ff1l Bfll l&trf* tibli Kesti territooriumil aitas pgdrapopulatsioonl r»l ka DOPÜlataloonl alatthiku (geograafшве, lokaal-. elemantaarpopulatsloonl) erist«^««уд- Terrltorlaaleete, erineva arvukuse ja populatsiooni struktuuri dflnsanlkaga populatsiooni alajactuate tekkimist ei vslealda Ilaselt põtrade suur vaghlsue ning rälke koha truudus, ais eri ranus- Ja soortihaadel on küll erlnsr. Lllkura "algrateioonipopulatslooni" suur osatähtsus real aastatel teeb rftlaatuks püsivate erinevustega piirkondade kujunemise. On tbenäoline, et kbigll aladel, kua psdral on eristatud lokaal- r6i e1ementaarpopulatsioone, pikema ajavahemiku jooksul andmeid kogudes erinevused kas kaovad vöi selgub, et need on tingitud tolturnis- v8i teiste tingimust* erinevusest ja et mitte oopulataloonl siseseid ajaühikuid vald antud asurkonda aoiutavate tegurite erineva intenaiivauaega alasid.isenditel, kee antud tingimustega ala piiridest lahkuvad ja satuvad teieta tingimustega alale, muutub toitumus ja siglmlee intensiirsus vastavaks antud tingimustele ning nad hakkavad sarnanema selle ala teistele isenditele; Järelikult hakkavad nad ka populatsiooni arvukuse dünaamika kujunemisel sarnanema seda ala varem asustanud isenditega. Ainsateks reaalselt erlstatavalks põtrade grupeeringulke on eri koapositsiooniga grupid. Hn. siglmlsgruppide (breeding groops), nagu neid arvatakse esinevat Alaskas /47/ Ja mis oleksid võrreldavad deeaiga /24/, "suurte perekondade" (Qroaa Familien /18/) v8i "partsellaarsete grupeeringute ehk populatsiooni partsellide" (парцеллярная группировка, популацжонная парцелла /167, 168, 169, 170/) dlemaeolu v8i puudumist meie materjal ei võimalda selgitada POPOLATSIOOrt STRUKTOUBID JA H0HÖ0STAA5 Populatsiooni tunnistamine keerukaks süsteemiks tähendab ka tunnistada, et populatsiooni spetsiifika ei ammendu teda moodustavate elementide iseärasustes, raid nende elementide rahaliste seoste ja suhete Iseloomus. Ble- 61
 populate!")
 struktuuri ühikud ja nende ökoloogilis-etoloogiline")
63 Т а Ъ a 1 52 Liigi kui süsteemi jaotuse printsiibid, jaotuse aluseks olevad seosed, alasüsteemid ja struktuurid Jaotuse printsiipi süstemaatiline ökoloogiline organisatsiooniline Jaotuse aluseks olevad seosed 1 Biosüsteemid: Alasüsteemid: geneetilised ökoloogiliseu etoloo fe iliseci liik /ix elamliik allopatrilised (territoriaalsed) populate!- liigi asurkond biflt^önoosis geograafiline pomi^tsioon okaalpopulatsioon eleaentaarpopulate ioon territoriaalne gru^e^rine aalsed) vanuseline struktuur sooline struktuur generatsiooniline struktuur konetituteionaalne struktuur tervislik struktuur kobporient Ж eltalnt 59 <0 э e 1з tabel 53 Põdrapopulatsiooni allopatrilise (territoriaalse) struktuuri ühikud ja nende ökoloogilis-etoloogiline iseloomustus Territoriaalsed ühikud ökoloogilis-etoloogiline iseloomustus liigi asurkond lligisisesed suhteu, lilbispetslifliine käitumine ; alsmllik lligisisesed suhted, geograafilisele populetsloonlle spetsiifilised käitumise iseärasused j populateloon lligisisesed suhted, im- ja emlgrantiue esinemine, osaline panmlksie; välisteyureist mõjutavad regionaalsed faktorid; territoriaalne grupeering de ей grupp komponent territooriumi jaotus grupeeringute vahel, uletuslik panmlksia,ajutine grupeeringute liitumine ja isendite (gruppide) vahetus; vällategureiet mõjutavad lokaalsed faktorid; hierarhilised suhted hierarhlliaed suhted sugupoolte vahelised suhted, sugulaslikud suhted, suhted Indiviidide vahel element - territooriumi ühis- 1 kasutus 62


64 mantide vahelisi seoseid ja suhteid võib vaadelda vähemalt kolmel tasandil - ruumilisel, ajalisel ja funktsionaalsel - ning katse anda "... ammendavat kirjeldust bioloogilisest objektist kui terviklikust süsteemist ühte tüüpi seose alusel, on teadlikult määratud ebaõnnestumisele. Probleemi lahendus seisneb täielikus ja vastuoludeta sünteetilises kõlgi nende seosetüüpide hõlmamises ja see tähendabki nsuet süsteemselt uurida," väidavad I.V. Blauberg ja E.G.Judin/101, lk. 168/. Nagu märgib V.D. Fjodorov /206/, v8ib formaalselt k8igi elementide ühiseks omaduseks pidada väga mitmet laadi informatsiooni vahetust. "Sel juhul, kui üks konkreetse informatsiooni vormidest..., saabunud ühelt isendilt, kasutatakse teise isendi poolt "otsuse vastuvõtmiseks" (käitumise, aktiivsuse reguleerimiseks), kujuneb nende vahel seos." Selline seos "informatsiooni alusel" pole muidugi ainuke seose tüüp, kuid nagu kõik teisedki tüübid, v6ib ka see esineda nii ruumilisel, ajalisel kui ka funktsionaalsel tasandil. Liigi kui süsteemi jaotusest, alasüsteemidest ja struktuuridest annab ülevaate tabel 52. Tebelist selgub ka, põhiline vastuolu populatsiooni msiate Ja ta alatlhikute piiritlemisel S.S. Schwarzi ja tema koolkonna ning N.P. Haumovi ja tema koolkonna vahel on tingitud sellest, et esimesed lähtuvad süstemaatilisest, teised aga ökoloogilisest jaotusprintsilbist. et Näib otstarbekas olevat liigi alasüsteemide ja vastavalt ka populatsiooni struktuuride jaotamine kahte suurde rühma ruumilisel tasandil: allopatrilised e. territoriaalsed ja sümpatrilised e. aterritoriaalsed alaeüeteemid ja struktuurid. Neid kahte rühma ühendab populatsiooni sotsiaalne struktuur. Põdra populatsiooni vastavate struktuuride ühikute ja elementide klassifikatsioon ning nende iseloomustus on edasi antud tabeleis 53 ja 54. Nagu populatsiooni struktuure ei saa vaadelda etaatilistena, vaid dtinaamilietena, nii ka ühe struktuuri piires seda moodustavad elemendid pole alati üheselt määratavad, vaid võivad muutuda sõltuvalt tingimustest» sama isend v8ib teatud tingimustel kuuluda kas ühte v8i teise elemendirühma. Näitena võib tuua põdra lehmmullika (1,5 a. vanune): soodsatel tingimustel kuulub ta populatsiooni sigivasse ossa, s-t. teda võib lugeda adultseks, halbadel tingimustel ta aga ei ьз


65 tabel 54 Põdrapopulatsiooni eiimpa trilised ( a territoriaalsed) struktuuri ühikud ning nende ieelooeuetub orfolooglline iseloomustus SoellM»traktuur ökoloogiline Iseloomustue aigimisvöimetud e ig laia võime liiased. popaleteiooni reproduteeeriv osa sigimisvõimel!^»ed. populetel-' odei reprodutsfeerlv "ose morfoloogiline Iseloomustus ;Ju veni Ilsed seniilsed noored keelte alleed vanad moored keskealised emad Vanuseline вtruituur ökolqog U im-e toloog il in«iseloomustus a ühtel Ise lt arvukae гшш, kuld auuze suremusega i tihedale sots. seoseis $$-* väheetrvullne, etolooglliae pärandi kandjad erinev jäzglaate erv, ahtruae %, vellategurelet eõ Ju tat we määr$ - etoloog 11 lee pärandi kandjad auur vaglilsus kõige elajöttllaemed - etolooglliae plxemdl kamdjad tabel 55 Populatsiooni omaduste hindaaieeks kasutatud näitajad ja generallseeritud hinnangud Populate looni osa iluaed. lodlkaator Oenerallaeerltud hlnnan* la/1000 ha aaoatubtihedus tlhedue proov la lal (ia/kv.) frekvente (%) produktiivaua tiheduse (frekvental) dünaa- juv. % ika territoriaalne «trak frekvents abundantsi tuur eoollne ja vanuseline etruktuuz aotalaalne etzaktuur Jaotuaue (ezi auuxuaega gruppide eeiaeaieaagedua) Jao toe ae (eri auuuaega gruppide koepoeiteloon) ia/1000 ha 6щ J Juv.sad. dtytjuv. 64


66 osale sigimisea ja kuulub populatsiooni mittesigivaese oeea, e. t. vasikate ja eubadultaete rühma. Loomulik on ka struktuurielemantide muutumine aj ae, muutumine teiseks atruktuurielemendike koos ksigi oma seoste ja suhetega. Seega populatsiooni struktuurid pole oma iseloomult mitte staatiline d, vaid dünaamilised ja vaetavalt ka populatsioon süsteem on tüüpiline dünaamiline süsteem. kui Kui vaadelda liigi kui süsteemi ja populatsiooni dünaamikat, е. o. neid nende muutuvuses, ajalisel tasandil, siis muutuste ulatus ssltub palju ruumist, kus tekivad vsi kaovad territoriaalsed ühikud ja vastavalt laieneb vöi aheneb liigi v8i populatsiooni poolt asustatav ala. Sel põhjusel ei saa populatsiooni (ja ka liigi) poolt asustatud ala vaadelda ühetaolisena, vaid perspektiivsuse ja kvaliteedi alusel mitmesse ossa Jagunevana s8ltuvalt populatsiooni seisundist ja konkreetseist populatsiooniväliseist tingimusist. Seega populatsiooni areaal jaguneb (1) Populatsiooni poolt asustatud alad. (1.1) Optimaalsete tingimustega alad (võimalik püsivalt suur asuetuetihedus). (1.2) Peeeimaaleete tingimustega alad (võimalik püsivalt madal asuetuetihedus ning lühiajalised asustustiheduse tõusud). (2) Populatsiooni poolt asustamata alad. (2.1) Blutingimustelt sobivad alad. (2.1.1) Optimaalsete tingimustega alad. (2.1.2) Peeeimaaleete tingimustega alad. (2.2) Blutingimustelt sobimatud alad. Populatsiooni seieundi hindamiseks mingil ajalsigul ja ta perspektiivide hindamiseks on eespool eaitatud populatsiooni areaali jaotuste silmaspidamine hädavajalik. Näiteks meie psdrapopulatsioonis lõppes aaatal populatsioonile elutingimuetelt sobivate alade hsivamlne (frekvents saavutas maksiaumi) ning kuna olid asustatud ka peeeimaaleete tingimuetega alad, algas järek optimaalsete tingimustega alade üleasustus (abundantsi hüppeline t8ue). Populatsiooni omadusi iseloomustavate näitajate generalieeerimise ohtudest ja kahjulikkusest oli juttu eespool (lk. 24), Populatsiooni eri omadusi täpsemalt kajastavatest

 käitumistüübile sõltuvalt konkreetsest situatsioonist väliskeskkonnas\".")
67 Indikaatoritest, mida kasutati autori poolt, Ja neile vastavatest generaliseeritud hinnanguist annab ülevaate tabel 55. Tuleb rõhutada, et kasutusele võetud näitajatest eriti tihedus, frekvents Ja jaotumus on populatsiooni produktiivsuse ja struktuuri detailsel analüüsil asendamatud. R. Roeeni /169/ jaotuse alusel kuuluvad populatsioonid adaptatiivsete süsteemide hulka, s. t. on süsteemid, "... mis vajaduse korral muudavad oma käitumist selliselt, et minna Ule eelistatumale (ehk optimaalsemale) käitumistüübile sõltuvalt konkreetsest situatsioonist väliskeskkonnas". Kuna sellise käitumise aluseks on stabiliseeriv regulatsioon, räägitaksegi ka populatsiooni puhul ta homöostaasist ja seda tagavatest homöoetaatilieteet mehhanismidest /139, 220/. Põdra- ja samade meetoditega uuritud metskitse-, metssea, valgejänese- ja rebaaepopulateiooni struktuuride nende dünaamika analüüs viis populatsiooni homöostaaei küsimuses järgmistele seisukohtadele. Populatsiooni homöostaasi ei tule mõista populatsiooni arvukuse taseme säilitamisena, vaid populatsiooni struktuuri teatud piirides hoidmisena. Populatsiooni arvukus on sekundaarne, struktuur aga primaarne. Muidugi mõjutab populatsiooni arvukus (asuetuetihedus) ka struktuuri, ja kuid enamail juhtudel oluliselt allee siis, kui arvukus (asuetuetihedus) antud elutingimustega alal ületab teatud piirväärtuse - tekib üleasustus - või kui arvukus (asuetuetihedus) on väikeem kui vajalik normaalselt eksisteeriva populateiooni struktuuri tagamiseka. Et populatsiooni omadused on oma iseloomult statistilised, sile ei saa ka populatsiooni homöostaasi mõista mingi antud liigile püsivate suurustega seisundina vaid seisundina, millel on oma varieeruvuse piirid, mille ulatuses seisund kvalitatiivselt ei muutu. Seega homöostaas iseloomustab populatsiooni kvalitatiivset seisundit ja see võib olla samasugune erinevate populatsiooni kvalitatiivsete näitajate puhul $ erineva arvukuse ja asustustiheduse, erineva populatsiooni vanuselise, soolise,"tervisliku jne. struktuuri puhul - nende näitajate muutuste teatud piirides. Sealjuures populatsiooni erinevad atruktuuriühikud võivad teatud määral üksteist kompenseerida või - vastupidise juhuna - üksteise puudusi progressiivselt suurendada. об


68 Populatsiooni homöoetaaei all tuleb mõista populatsiooni sellist seisundit, mis võimaldab tal eksisteerida, s.o. toota vajalikul hulgal järglasi ning säilitada eksisteerimiseks vajalikes piirides populatsiooni struktuur. Populatsiooni arvukuse, asustustiheduse ja levila lühiajaline ahenemine ebasoodsate tingimuste mõjul ei tahenda veel populatsiooni homöostaasi rikkumist: kui säilib populatsiooni struktuur, on endise taseme taastamine võimalik enamasti antud liigi ühe-kahe elutsükli jooksul. Kuna populateioonivälised tingimused on pidevas muutumises, muutub pidevalt ka populatsiooni seisund, eri etruktuurielementide kvalitatiivsed suhted, mis tingibki struktuuri taastavate homöostaatiliete mehhanismide rakendumise. Kuna nii välistingimuetel kui ka nende mõjul rakenduvatel homöostaatilistel mehhanismidel on oma inerts, pole kunagi täit vastavust populatsiooni momendi seisundi ja sel ajal rakendunud homöostaatiliste mehhanismide vahel. Seega populatsiooni normaalseks seisundiks on dünaamiline, mitte staatiline seisund ning populatsiooni homöostaasi tagend.eo aluseks on rütm, в. t. erinevate näitajate teatud sagedusega kõikumine, järkjärguline lähenemine antud tingimustes optimaalsele tasemele ja kavakindel taandumine elutingimuste muutumise korral - dünaamiline populatsiooni struktuuri Umberkorraldumine vaetavalt tingimuste muutumisele. Näib, et ökoloogilise homöostaasi puhul, vastandina geneetilisele ja füsioloogilisele homöoetaasile, on otstarbekae rääkida mitte ühest, vaid kolmest homöostaasi seisundist vastavalt populatsiooni eri seisundeiles 1) stabiliseerunud populatsioonis, kus elutingimustelt sobivad alad on kõik hõivatud ja' asuetuetihedus on optimaalsel (kulutatavate toiduressursside taastumisele võrdeel) tasemel, - normaalne homöostaae. mille puhul homöostaatilieed mehhanismid püüavad tagada produktiivsuse võrd- Bust suremuse pluss emigratsiooniga. Enamail juhtudel siin juhtivaks tagasiside asustustiheduselt sündimusele ja populatsiooni struktuuri ning produktiivsust määravad populatbioonisiseeed faktorid; juhul, kui emigratsioon ei taga populatsiooni produktiivsuse võrdsust suremusega, rakendub kaitsev homöostaae; b7 on

 populataioonis, kus olemasolevad elutingimused ja sobivad alad on üleasustatud (toitutervitatakse rohkem, kui taastub; ei Jätku liigi nsudlustele vaetaval määral territooriumi) - kaitsev")
69 2) populatsiooniв, nie pole veel saavutanud stabiilsust ja millel on küllaldaselt optimaalsete tingimustega asustamata eluruumi - ekspansiivne homöostaas, mille puhul homöoataasi mehhanismid on suunatud maksimaalse produktiivsuse tagamisele! iseloomulikuks on keskmise vanueo suurenemine (seosee vanemate populateiooniв emasloomade suurema viljakusega), varasem suguküpaue, suurem järglaste arv, emasloomade osa suurenemine jne.; 3) populataioonis, kus olemasolevad elutingimused ja sobivad alad on üleasustatud (toitutervitatakse rohkem, kui taastub; ei Jätku liigi nsudlustele vaetaval määral territooriumi) - kaitsev homöostaas. mille puhul homöobtaatilieed mehhanismid on suunatud populatsiooni produktiivsuse ja ta struktuuri sellisele muutmisele, et viia populatsiooni produktiivsus väiksemaks kui suremus plues emigratsioon. Iseloomulik on väga intensiivne emigratsioon, isaste osa suurenemine populatsioonis, sündimuse järsk vähenemine nii populatsiooni väliete kui ka sisemiste tegurite msjul ja tüüpilisena stressinähtub. Vaadeldes eespool toodud taustal meie psdrapopulatsiooni viimastel aastakümnetel, võime nentida, et kuni aastani oli populatsioon ekspansiivse homöostaasi seisundis; elutingimuste Järsk halvenemine (karmid talved, halvad suvised toitumistingimused) viisid aastail kaitsva homöoataasi rakendumisele, mis aastal asendus normaalse homöostaasiga ning seejärel ilmnesid uuesti esimesed käitava homöostaasi tunnused, tingituna asustustiheduse tunduvast suurenemisest PÕDRAPOPÜLATSIOOHI RATSIONAALSEST KASUTAMISEST Populatsiooni juhtimine ja ta ratsionaalne kasutamine tähendavad eelkõige populatsiooni struktuuri, arvukuse, leviku ja produktiivsuse teadlikku suunamist. On aegunud selle "juhtimise" all mõista ainult küttimise intensiivsuse muutmist. Kuna populatsiooni mõjutavad üheaegselt paljud tegurid ja nende mõju võib populatsiooni homöostaasi mehhanismidele olla kumulatiivne ja eri populatsiooni struk- 68

.")
70 tuuride suhtes valikuline, siis nsuab populatsiooni juhtimine populatsiooni eri struktuuride suunatud.1a kooskõlastatud asjutaaist, s. t. teadlikku populatsiooni eri struktuuride kvalitatiivsete suhete säilitamist vsi suunatud muutmist. Selleks on aga vajalik tunda eri struktuuride osa populatsiooni homöostaasi eri seisundeis, struktuuride muutlikkuse piire ja nende omavahelisi kvantitatiivseid seoseid Ja suhteid. Mingi loomaliigi kasutamise ulatuse määrab olemasoleva ja nn. lubatava asustustiheduse vahe igas konkreetses piirkonnas (jahimajandis). Kuna lubatava asustustiheduse ülempiir sbltub konkreetsetest tingimustest (looduslikest, eri majandusharude huvidest, antud ala looduslike tingimuste kasutamise eesmärkidest Jne.),ei saa see olla ühesugune suurtel territooriumidel ega ka määramata aja jooksult vastavalt tingimuste ja eesmärkide muutumisele muutub ka lubatava asustustiheduse ülempiir. Lähtudes psdra bioloogiast ja ta metsamajandusliku tähtsuse hinnangust /48, 51, 149/, loeti meie alal maksimaalseks lubatavaks asustustiheduseks isendit 1000 ha psdrale elutingimuetelt sobiva territooriumi kohta. Jahimaade boniteerimise ja korralduse käigus selgitati Jahipiirkondade lõikes eri boniteediga alade esinemise ulatus ja arvutati nende andmete alusel igas piirkonnas lubatav isendite arv. Kogu meie territooriumil vbiks nendel andmetel psdra p&hipopulatsiooni kevadine arvukus olla isendit, s. t aasta arvukusest umbes kaks korda väiksem. Metsamajanduslike kahjustuste pidamine vähemoluliseks põdrast saadavast tulust on tinginud liiga madalate laskenoimide määramise, mis pole kaasa aidanud psdra arvukuse madaldamisele lubatava asustustiheduseni. Sellega on kaasnenud populatsiooni viljakuse vähenemine, mitmete sotsiaalse struktuuri häirumiet iseloomustavate indikaatorite /10/ ilmumine, nn. hädatoidu (kask, kuusk, oras jm.) sage tarvitamine ning v8ib olla ka üldine kasvu ja kehakaalu vähenemine (vt. teised tööd samas kogumikus). Populatsiooni sisemiste v&imaluete maksimaalseks rakendumiseks produktiivsuse suurendamisel on otstarbekas hoida populatsiooni ekspansiivse homöostaasi seisundis, s.t. hoida asuetuetihedus optimaalsest tasemest madalamal küt 69
 5 g В E X -kl -1 etruktuur.")

71 timise vaetava intensiivsusega ning eamal ajal säilitada ekspansiivse homöoataaei eeieundile vaetav populatsiooni T a b a 1 56 Põdrapopula ts iooni perspektiivne produktiivsus (vasikate % sügisel) olenevalt eelnenud talvede-suvede tingimustest ( aastate analüüsi andmeil) 5 g В E X -kl -1 etruktuur. Selle eeemärgi tagamieeke on vajalik viia populat eiooni arvukus lubatava asustustiheduse ja arvukuse tasemele ning hoida ta eellel tasemel Bige laskenormi rakendamisega, eäilitadee samal ajal ka populatsiooni Bige struktuuri. Vasikate arv s-š Д i Ji Jj 0^0 1.. n H 0 J3 d ^0 il НИ а ai 1 ет 60,4 И. M juv. 2Juv 25 E > VI R ОН * * > M *+ 4> * * +4- * 60 Ч-4-50 H > 0 4-4» 4» M Tühistus 1 - pehme talv, SüOOIlC suvi 2 - kaim talv, ebasoodne +>4- - suur ++ - keskmine 0 - puuaub Õige laskenormi määramisel, s. o. eelle prognoosimisel, on kbige olulisemaks ette näha populatsiooni produktiivsuse muutusi sõltuvalt populatsiooni struktuurist ja looduslikest tingimustest. Palju vbib siin kaasa aidata populatsiooni arvukuse dunaamika modelleerimine /31, 35, 36, 204/, kuid ainult tingimusel, et modelleerimisel arvestatakse võimalikult täpselt kbiki produktiivsust mõjutavaid tegureid nende dünaamikas. Hiljuti V.P. Galantsevijt./108/ poolt tehti katse matemaatiliselt modelleerida Leningradi pbdrapopulatsiooni dünaamikat. Selle pbhipuudusteks on populatsiooni dünaamikat mõjutavate faktorite pidamine kas 70


72 staatilisteks v6i siis pika aja jooksul ühtlaselt muutuvaike, maksimaalselt generaliseeritud andmete kasutamine, populatsiooni pidamine suletud süsteemiks samal ajal, kui tegu on lahtise süsteemiga jne. Tõeliselt populatsiooni dünaamikat prognoosida võimaldava mudeli puudumiseni on v8imalik juhinduda käesolevas töös populatsiooni struktuuri ja produktiivsuse analüüsil saadud tulemustest, mille üldistus on esitatud tabelis 56. Hoopis raskem on Õlge laskestruktuuri tagamine, kuna suur laekenorm ja kuulipüsside vähesus ei vbimalda meil kuigi tagajärjekalt teha valiklaskmist. Väga perspektiivne oleks meil rakendada S.S. Schwarzi poolt korduvalt soovitatud nn. ökoloogilist vaakumit /218, 222/. Ökoloogilise vaakumi mõte seisneb selles, et populatsiooni areaalis huakse tühikud, kust antud liigi isendid kõrvaldatakse. Sellise ala asustamisel kujuneb seal iseendast 8ige struktuuriga populatsioon ekspansiivse homöostaasi seisundis. Sar mai ajal sellise vaakumi kujundamisel tuleb vaetaval alal küttida k8ik antud liigi isendid ning järelikult ei rikuta populatsiooni struktuuri, kuna see väljavõte pole valikuline. Arvestades psdra suurt liikuvust, vsiks ökoloogilise vaakumi ala hslmata % talle elutingimustelt sobivast alast ning paikneda migratsiooniteedel, millega saavutatakse populatsiooni migreeruva ja liikuvama osa intensiivsem kasutamine võrreldes paikee osaga. Küttimise kontsentreerumine ainult kuni ühele viiendikule alale vähendaks ka emotsionaalse stressi ebasobivat mõju ning aitaks kogu meie alal uuesti kujundada ekspansiivse homöoataasi seisundis oleva populatsiooni. 71
 et populatsiooni struktuur ja temas toimuvad protsessid on väga")
 kuna populatsioon on keerukas bioloogiline süsteem, siis on selle tu")
73 K o k k u v õ t e Populatsiooni struktuuri süvendatud uurimisel lähtuti järgnevast» 1) kuna populatsiooni struktuur ja temas toimuvad protsessid on oma olemuselt statistilised nähtused, on vajalik andmete statistiline analüüs ning meetodite rakendamine, mis võimaldavad saada statistilise analüüsi nõuetele vastavaid materjale; 2) et populatsiooni struktuur ja temas toimuvad protsessid on väga dünaamilised nii ajas kui ka ruumis ning neid mõjutavad paljud muutuvad tegurid, on populatsiooni kui terviku ja ta üksikelementide muutuste tegelike piiride ning omavaheliste seoste selgitamiseks vajalik koguda andmeid pikema ajavahemiku jooksul; 3) kuna populatsioon on keerukas bioloogiline süsteem, siis on selle tundmaõppimiseks vajalik süsteemne lähenemine ja süsteemne analüüs, s. t. tuleb selgitada populatsiooni alasüsteemlde kvantitatiivne koostis ning nendevaheliste seoste iseloom ja tugevus nende ajalises ja ruumilises muutuvuses. Põdra populatsiooni sigimise ning 1 ta soolise, vanuselise, sotsiaalse ja territoriaalse struktuuri analüüsimiseks kasutati aastate sügie-talvistel ajujahtidel kogutud andmeid ajus olnud Isendite arvust, nende soost ja vanusest (kas vasikas või vanem). Kokku laekus 11 aasta jooksul andmeid ajudest kvartalis, millest aeti välja põdragruppi kokku Isendiga. KÕlkl andmeid analüüsiti igal aastal viie analuüsirajoonl ja kolme kuu lõikes. Koos vaadeldi ainult andmeid, mis usaldataval tasemel ei erinenud. Eri aastate, eri analüüeirajoonide ja eri kuude andmete erinevuste-eamasuete kind- 72


74 na-eeetie. Kogu vaadeldava perioodi oktoobrikuu andmete võrdlusel analüüsirajoonide lõikes Ilmnes, et eel ajavahemikul esines kokku viis omavahel oluliselt erinevat jaotumustüüpi, mille ajalises ja ruumilises levikus esinevad kindlad seaduspärasused! kahte neist jaotumustütlpidest v8ib pidada põhilisteks, kuna üke on tileminekutüüp ja kaks tüüpi on kujunenud populatsioonleieeste liikumisete tulemusena ning on seega ka nende liikumiste indikaatoriks. Võrdlus naaberaladega näitab, et aastail langes Leningradi oblastile Iseloomulik põdrakarjade jaotumus usaldatavalt kokku Ida-Bestis (Alutagusel) täheldatuga. Jaotumast määravaist tegureist on olulisemaiks klimaatilised tingimused; populatsiooni arvukus (asuetuetihedus) mõjutab jaotumuet alles teatud tiheduse piiri ületamisel (peale elutingimustelt sobivate alade asustamist); oluliselt mõjutavad jaotumuet populatsioonieieesed liikumised (jaotumuste analüüs võimaldab nende esinemist ja ka liikumiste suunda selgitada!) ja sotsiaalne struktuur; võrdlus Poissonl jaotusega näitas põdrakarjade jaotumuse usaldatavat erinemist kõigil kuudel, aastatel ja kõigis analüüsirajoonldes. Arvestades Eestis kogutud andmeid põdrapopulatsiooni sigimise ja struktuuride dünaamikast, ei saa populatsioonisiseete ühikute eristamise aluseks võtta erinevusi produktiivsuses või populatsiooni struktuurides, nagu seda on tehtud. Meie materjali analüüs näitab, et pole mingit alust Eesti territooriumil eristada mitut põdrapopulatsiooni või populatsioonisisest alatihikut (geograafilist, lokaal- elementaarpopulatslooni). Populatsiooni 8traktuuri ja produktiivsuse või näitajana kasutatakse sageli liialt generalieeeritud hinnanguid, mis moonutavad tegelikkust ega võimalda leida eri näitajate vahelisi seoseid ega selgitada struktuuri, produktiivsust ja nende dünaamikat suunavaid tegureid. Populatsiooni struktuuri ja produktiivsuse detailanalüüeil on aeendamatuiks sellised indikaatorid nagu tihedus, frekvents, jaotumus, kaksikutega emaloomade osa vasikaga emaloomadeat jt. Populatsiooni homöoataasi tuleb ökoloogilisest aspektist vaadelda mitte populatsiooni arvukuse taseme säilitamisena, vaid populatsiooni struktuuri hoidmisena. Vastavalt populatsiooni eeisundeile on eristatud ka kolm homöostaasi 75
75 seisundit: normaalne, ekspansiivne ja kaitsev homöostaae. Populatsiooni ratsionaalseks kasutamiseks on vaja põtrade arv viia metsamajanduslikult lubatavale tasemele ning hoida õiget laskenormi ja -struktuuri rakendades ekspansiivse homöostaasi seisundis. Bige laskenormi määramist võimaldab populatsiooni produktiivsuse prognoosimine üldistava skeemi abil, õige laskeatruktuuri tagamiseks on ettepanek rakendada S.S. Schwarz! poolt soovitatud "ökoloogilist vaakumit". 76

. - XI Intern. Congr. of Came Biologiste. Stockholm, 1974, p. 59-66. 6.")
76 K i r j a n d u s 1. A l l e n, D. L. O f f i r e, m o o s e a n d w o l v e s. - " A u d u bon", 1974, Vol. 76, Ho. 6, p A u l, J., L i n g, H., P a a v e r, K. E e s t i. H S V imetajad. Tln., 1957, lk B a l l e n b e r g h e, V., P e e к, J. R a d i o - telemetry studies of шооае in northeaeten Minnesota. - "J. Wildlife Manag.", 1971, Vol. 35, Но. 1, p B e r g e r u d, А. Т. 8 u m b e r e a n d d e n s i t i e s.- I B P handbook Ho. 7, 1969, p B l o o d, D. A. V a r i a t i o n i n r e p r o d u c t i o n a n d p r o duction and productivity of an enclosed herd of mooee (Alcee alcee). - XI Intern. Congr. of Came Biologiste. Stockholm, 1974, p B o n a c k. A. W e s t w a n d e r u n g v o n E l c h e n, A l c e e a l cee. - "Säugetierkundl. Mitt.", 1961, Bd. 9, Hr. 4, S B r i e d e r m a n n. L. D i e W e s t v o r s t ö ß e d e e E l c h wildes (Alcee alcee L.) in Mitteleuropa in ihrer populationsdynamischen Bedeutung. - Zool. Garten H. P.,1968, Bd. 35, Hf. 4/5, S B u b e n i k. A. G r u n d l a g e n d e r W i l d e r n ä h r u n g. Berlin, Deutscher Bauernverlag, 1959, S В u b e n i к, A.B. Probleme der Wilddichte im Lichte der Wildsoziologie. - "Schweizerische Zeitschrift fur Forstwesen", 1967, Bd. 118, Hr. 2, S Bubenik, A.B. North American moose management in light of European experiences. - 8 th Horth 77


77 American Moose Conference and Workshop. Thunder Bay, Ontario, 1972, p , 19, 20, 21, C h a p m a n, P o p u l a t i o n e s t i m a t i o n b a s e d cange of composition caused by a selective removal. - "Biometrika", 1955, p C u r r y - L i n d a h l, K. A n o t e o n t h e m o o s e i n Sweden. - "Terre et vie", 1961, Vol. 108, No. 1, p D a h l b e r g, B. L. T h e m o o s e r e t u r n s. - " W i s c o n sin Conservat. Bull.", 1964, Vol. 29, No. 3. p D a n i e l, 0. J o o n i j a h i n d u s e a l a l t. I I. - " E e s t i mets", 1934, lk D о d d s, D.G. Observations of pre-rutting behaviour In Newfoundland moose. - "J. of Mammalogy", 1958, Vol. 39, No. 3, p E b e r h a r t, L. P o p u l a t i o n a n a l y s i s. - W i l d l i f e Management Techniques, Washington, 1971, p E d w a r d s, R. Y., o n R i t с e у, R. W. R e p r o d u c t i o n in a moose population. - "J. Wildlife Manag.", 1958, Vol. 22 (3), p E i b l - E i b e s f e l d t, I. B e i t r ä g e z u r B i o logie der Haus- und der Ährenmaus nebst einigen Beobachtungen an anderen Nagern. - "Z. Tierpsychol.", Berlin, 1950, Bd. 7, S E i s e n b e r g, J. P. T h e s o c i a l o r g a n i z a t i o n s o f Mammals. - Handbuch der Zoologie, Berlin, 1966, Bd. 8, S P i 1 о n о v, C.P., Z у к о v, C.D. Dynamics moose populations in the forest zone of the European part of the USSR and in the Urals. "Natur, can.", 1974, Vol. 101, No. 3-4, p P r e n z e 1, L.D. Occurence of moose in food wolves as revealed by scat analyses: a rewiew of North American stydies. - "Natur, can.",1971ц Vol. 101, No. 3-4, p of of


78 22. G а в a w а у, W.A., С о a d у, J Л. Review «f energy requrements and rumen fermentation in mooее and other ruminants. - "Hatur. can.", 1974, Vol.101, Ho. 1-2, p G e i s t, V. A behavioural approach to the management of wild ungulatee. - "Sei. Manag. Anim. Plant Communit. Coneerv.", Oxford, 1971, p G i 1 m о u r, J.S.L., G r e g о r, J.W. Demes: a suggested new terminology. - "Hature", 1939«Vol laketegemieeks kasutati tavaliselt ^-menetlust, samuti kasutati seda jaotumuste omavahelisel võrdlusel ja v8rdlueel Poissoni jaotusega. Seoste ja nende tugevuse selgitamiseks kasutati korrelatsioonikordajat r ja astakkorrelatsioonikordajat r g. Real juhtudel arvutati korrelateioonimaatriksld raalil "Minsk-32". Populatsiooni sigimise ja produktiivsuse selgitamisel kasutati kümmet eri näitajat, mille dünaamika ja omavaheliste seoste analüüs ning nende seostamine konkreetsete looduslike tingimustega võimaldas püstitada rea sigimisnäitajate dünaamika tüüpe, mida saab rakendada populatsiooni produktiivsuse prognoosimisel. Populatsiooni produktiivsuse alusel eristati kolm perioodi - kaks kõrge viljakusega perioodi ja nende vahel madala viljakuse periood. Kolmandal perioodil saavutati esimese perioodiga peaaegu võrdne produktiivsuse tase, kuid samal ajal oli vähenenud kaksikutega emaloomade osa ja suurenenud sigimisest osavõtvate ja ühe vasikaga emaloomade osa. Põhjuseks on kas emasloomade vanuselise struktuuri muutus (küttimine, immigratsioon) vm eelnenud ebasobivate tingimuste mõjul nõrgema konstitutsiooniga aastakäikude suurem osa sigivate emasloomade hulgas. Vastavate andmete analüüs kogu põdra areaali ulatuses näitas, et potentsiaalne viljakus on põdral kogu areaali ulatuses sama, selle realiseerimine aga sõltub konkreetsetest tingimustest. Näiteks Eestis esines vaadeldaval ajavahemikul aastaid, kus produktiivsus saavutas kogu areaali ulatuses täheldatuet nii maksimaalse kui ka minimaalse suuruse. Kolmikute-nelikute esinemissageduse ja kirjanduses toodud andmete analüüs võimaldas järeldada, et nende esinemine pole mitte juhuslik ja haruldane nähtus (ei vasta Poissoni jaotusele!), vaid seaduspärane, mis võimaldab kolmikute-ne-


79 3 3» H o u s t o n, D. A s p e c t e o f t h e s o c i a l o r g a n i s a t i o n of moose. - IUCI Publ. Hew eer., 1974, Vol. 1, Ho. 23/2, p Idetroa, J.M. The Moose in Minnesota. - Techn. Bull. Minn. Dept. 1965, Ho. 9, P uonserv. Pish and вате, 35. J о г d a n, P.A., В о t к i n, D.B., Tolfe, M.L. Bicease dynamics in a moose population. "Ecology", 1970, Vol. 52, Ho. 1, p J u n i o r, P. M a t e m a t i e c h e O e d a n k e n U b e r d i e a l - tereklassenmäßig gegliederte Bewirtschaftung einiger Schalenwildarten. - "Z. Jagdwiss.", 1966, 12, Hr. 4, S К a e m p f, T. Obserwacje nad lo si ami Puezczy Kaapinoskiej. - "Lowiec polski", 1966, nr. 1, К в 1 к е г, G.H. Estimating Deer Populations by a Differential Hunting Lose in the Sexes. - Proc. Utah Acad. Sei., Arts and Letters, 1940, Vol. 17, p К e 1 к e r, G.H. Sex-ratio equations and formulae for determining wildlife populations. - Proc. Utah Acad. Sei., Arte and Letters, 1940, Vol. 19, p Kelsall, J. Structural adaptations of moo ее and deer for snow. - "J. of Mammalogy*, 1969, Vol. 50, Ho. 2, p ( о i r i в t о, I. Hirvikantamme rakenteeeta, lieääntymieeeta Ja verotukseeta. - "Suomen Riista", 16, 196З, в Koivieto,!. Hirvi. - Suomen nieakäät, 2. Helsinki, 1972, в К r a f f t, A. Management of moo ее in a norwegian forest. - "Medd. Stat, viltundereok.", 1964, Ser. 2, Ho. 16, p К u m a r 1, Б. Eeeti HSV linnud. Tln., Lent, P.C. A rewiew of rutting behavior in mooee. - "Hatur. can.", 1974, Vol. 101, Ho. 1-2, p
 Ja metsamajandus. - 2BSV TA Looduseuurijate Seltsi aastaraamat, 54. kd., Tartu, 1961, lk. 227-249. 49.")
 Population in the Estonian SSR. - \"Suomen Riista\", 1970, Bo. 2, p. 14-23. 51. L i n g, H.")
80 46. LeResche.R. Migrations and population mixing of moose on the Kenai Peninsula (Alaska). In: 8 th Borth American Moose Conference Workshop, Ontario, 1972, p and 47. LeResche, R.8. Moose migrations in Borth America. - "Hatur. can.", 1974, Vol. 101, Bo. 1-2, p L i n g, H. Põder (Alcea alcee L.) Ja metsamajandus. - 2BSV TA Looduseuurijate Seltsi aastaraamat, 54. kd., Tartu, 1961, lk L i n g, H. P8dra minevikust, olevikust ja tulevikust. - "Eesti Loodus", 1962, nr. 6, lk L i n g, H. On the Composition and Productivity of the Moose (Alcee alcee) Population in the Estonian SSR. - "Suomen Riista", 1970, Bo. 2, p L i n g, H. On productivity dynamics of the Betonton moose population. - "Aquilo Ser. Zool1972, Bo. 13, p L i n g, H. Veel kord põdra minevikust, olevikust ja tulevikust. - "Eesti Loodus", 1972, Br. 10, lk «L i n g, H. On the possibilities of employing abundance, frequency and density in sample areas for estimating numbers of moose (Alcee alces L.). - Estonian Contr. to the Intern. Progr. VIII. Tartu, 1975,p Biol. 54. Markgren.G. Puberty, dentition and weight of yearling moose in a Swedish country. - "Viltrevy", 1964, Vol. 2, Ho. 7, p Markgren.G. Reproduction of moose in Sweden. - "Viltrevy", 1969, Vol. 6, Ho. 3, p Markgren.G. The mooee in Fennoecandia. "Hatur. can.", 1974, Vol. 101, Ho. 1-2, p II 81


81 57. Markgren.G. Factors affecting the reproductions of aooee (Aloes alcee) in three different Swedish areas. - XI Intern. Congr. Game Biologists. Stockholm, 1974» p Markgren.G. The question of polygamy at an unbalanced sex ratio in the moose. - Intern. Symp. "The behaviour of ungulates and its relation to management*. Dniv. of Calgary. Switzerland, 1974, p Marteneon, A. Wild, Wald und Jagd in den Ruse. - Ostseeprovlnzen. Beudens», McGillis.J.R. The Kidney fat index as an indicator of condition in various age and sex classes of moose. - Ins 8 th Sort American Moose Conference and Workshop, Ontario, 1972, p Mereste, ü. Statistika üldteooria. Tln., 1975* lk Mivkwitz.A. Zwei Bicharten? - "Baltische Waidmanneblätter", 1901, S Mysterud,!. Behaviour of the brown bear (Ureus arotoe) at moose kills. - "Horweigian Journal of Zoology", 1973, Vol. 21, Ho. 3» p Mäkin.en,Y. Tilastotiedettä biologeille. Turku, 1974, e Nolcken,A. Zwei Elcharten. - "Baltische Waidmannsblätter", 1901, S Novakova,E., Hanzl,R. Standortebedingungen und Hirschgeweihqualität. - "Schweiz. Z. Forstwesen", 1967, 118, Nr. 10, S Oldemeyer, J.L. Nutritive value of moose forage. - "Natur, can.", 1974, Vol. 101, Nr. 1-2, p Onderscheka.K. Wildtierernährung und Wüdtierökologie aus der Veterinärmedizin.-"Allg. Forstzeitung", 1975, Bd. 86, Nr. 2, S of


82 69. P a г 1 к, t). Jahipidamine ебганetele Beetle. Jaht Ja ulukid", EH SV Jahlmeeete Selt ei aastaraamat , Tln., 1974, lk Peek, J.H. A review of moose food habite studies in Borth America.- "Hatur. can. ",1974, Vol. 101, Ho. 1-2, p P e e к, J., L о v а а в, A. Diferential distribution of elk by вех and age on the Gallatin winter range Montana. - "J. Wildlife Manag.", 1968, Vol. 32, Ho. 3, p Petereon, R.L. Horth American moose. - Toronto, Petereon, B.O., Allen, D.L. Snow conditions ae a parameter in mooeewolf relationships. - "Hatur. can.", 1974, Vol. 101, Ho. 3-4, p Phillipe, R.L., Berg, W.E., S i n i f f, D.B. Moose movement patterns and range use in Horth-Weetern Minnesota. - "J. Wildlife Manag; 1, 197З, Vol. 37, Ho. 3, p Pieloweki.Z. Die Wied ereinbtlrge rang dee Slcb.es-Alcee aloes (L.) im Kamplnoe-Hatlonalpark in Polen. - "Z. Jagdwlse?, 1969, Bd.15, Hr. 1, S Pimlott, D.H. Reproduction and productivity of Hewfoundland moose. - "J. Wildlife Maring", 1959, Vol. 23, p Pulliainen, E. Süden ravinnoeta ja saali stuetavoieta. - "Suomen Riista", 1963, Nr. 16, s Pulllalnen, E. Studies on the wolf (Canie lupus L.) in Finland. - Annal. Zool. Fennici, 1965, 2, p «Pulliainen, E. Seasonal movements of moose in Europe. - "Natur, can.", 1974, Vol. 101, No. 1-2, p


83 80. Ei II о t, )i Haruldasemate et jahiloomade et Beetle (Surieed reisimuljed Virumaa ja Tartumaa metsadest.). - "Beeti Kütt", 1922, lk Rajakoski,B., Koivisto.L. Hirvinaaraiden eukukypeyysiästä Suomeeea. - "Suomen Hiieta", 1966, 18, s Rausch, H.A., Brathie, A. Annual of moose calf production and mortality in southcentral Alaska. - In: Proc. 45 th Annual Cauf. West Assoc. State Game and Pish Commissioners, Auohorage, Alaska, 1965, p S a s e, P. Zum Artikel des Herrn К. Greve, Moskau, Zur Geschichte der Ausrottung des Elches Europa, und seine heutige Verbreitung. - "Baltische Waidmanneblätter", Schladweiler,F., Sterens,D. Reproduction of ehiras moose in Montana. - In "J. Wildlife Manag.", 1973, Vol. 37, Но. 4, p Schwerdtfeger.P. Ökologie der tlere.bd. 2. Demökologie. Hamburg und Berlin, 1968.' 86. S i m к 1 n, D.W. Reproduction and productivity of moose Northwestern Ontario. - "J. Wildlife Manag.", 1965, Vol. 29, p S 1 m к 1 n, D.W. Reproduction and productivity of mooee. - "Natur, can.", 1974, Vol. 101, No. 3-4, p S к u n с e, P. Algen. Studier, Jakt och värd. Stockholm, Stringham, S.P. Mother-infant relatione in moose. - "Natur, can.", 1974, Vol. 101, No. 1-2, p V e r m e, L.J. Reproductive patterns of whitetailed deer related to nutritional plane. - "J. of Wildlife Manag.", 1969, Vol. 33, p V о в, А., В г о x,, P., Geiet.V. A Reiew of social behaviour of the North American " 4

, 15, 3, с. 200-202. 97. Аршавский И.А. К обоснованию понятия \"функция развития\" во внутриутробном периоде млекопитающих и человека в связи с проблемой профилактики физиологической незрелости.")
84 Servide during the reproductive period. Inj The American Midland naturalist. The University of Hotre Dame, Hotre Dane, Indiana, 1967, 7ol. 77, Жо. 2, p Waemuth,P. Tabellarische Haturgeschichte der Säugetiere d. Oetseeprovinsen. Reial, W e b e r, В. Grundrias der Biologlechen Statistik. Jena, Gustav Fischer Verl., 1967* 94. W о 1 f, J. Einiges über unser Elchwild.-"Baltische Waidmannsblätter", 1902, S Wolfe, M.L. An overview of moose ooactions with other animals. - "Hatur. can.", 1974, Vol. Ю1, Ho. 3-4, p Андреенко К.П.. Стрельников А.П.. Тихонов И.Ф.К вопросу изучения причин облысения лосей. - "Уч. зап. Петрозаводского ун-та", 1967 (1969), 15, 3, с Аршавский И.А. К обоснованию понятия "функция развития" во внутриутробном периоде млекопитающих и человека в связи с проблемой профилактики физиологической незрелости. - В сб. "Проблемы современной эмбриологии", Изд. Ленинградского ун-та, 1956, с Балейшис P.M. Определение наступления половой зрелости Alces alcee L. по слоям дентина резцов. - "Первый междунар. териолог, конгресс", М., 1974, с Банников А.Г., Теплов В.П. Движение численности и плотности населения лося в РСФСР. - В сб. "Биол. и промысел лося", I, М., Россельхозиздат, 1964, с Беили Н. Статистические методы в биологии. М., Изд. иностр. лит., 1962, с Ьлауберг И.В., Юдин Б.Г. Становление и сущность системного подхода. М., "Наука", 1973, с Ролденков С.В. Динамика численности и некоторые данные о добыче лося в украинской ССР. - В сб. "Развитие охотничьего хозяйства украинской ССР", Киев, 1973, с Бочаров М.К. Методы математической статистики в геограгии. J., "Мысль" 1971, с


85 104. Бубеник А.Б. Плотность населения охотничьи животных, кормовая емкость угодий и повреждение леса охотничьими животными. - В сб. "Биол. и промысел лося", * 2, М., Россельхозиздат, 1965, с Василевич В.И. Статистические методы в геоботанике. Л., "Наука", 1969, с Верещагин Н. Лоси в Ленинградской области. - "Охота и охотн. х-во", 1965,» II, с Галака Б.А. Лоси пришли в Карпаты. - "Природа", 1969, Ш 12, с Галанцев В.П., Кисдяков Ю.Я., Скалинов С.В. Математическое моделирование динамики популяции лося (Alcee aloes). - "Зоол. ж.", 1975, т. 54, вып. 5, с Гептнер В.Г. Некоторые черты истории и современное состояние ареала лося в европейской части Советского Союза. - В сб. "Вопр. охотн. х-ва СССР", М., "Колос", 1965, с ПО. Гептнер В.Г., Насимович A.A., Банников А.Т. Млекопитающие Советского Союза. Т. I. М., "Высшая школа", 1961, с Герасимов Ю.А. Опыт авиаучета лосей в Московской области и проверка результата методом математической статистики. - В сб. "Совещание по вопросам организации и методам учета ресурсов фауны наземных позвоночных". Тезисы докладов, М., Гимельфарб A.A., Гинзбург Л.Р., Полуэктов P.A., Пых Ю.А.. Ратнер В.А. Динамическая теория биологических популяций. - Под редакцией P.A. Лолуэктова. М., "Наука", 1974, с ИЗ. Горегляд Х.С., Литвинов В.Ф. Болезни лосят. - В сб."березинский заповедник", вып. 3, Минск, "Урацжай", 1974, с Дементьев В.И. Численность и структура популяции лосей Ленинградской области. - "Материалы Научно-техн. конференции Лесохоз. фак. Ленингр. лесотехн. акад. 1967", Л., Джупина С.И., Свиридов A.A. Ящур в условиях эксперимента. - "Ветеринария", 1965, К' 5, с


86 116. Джурович В.М., Михайлов А.П. Опыт одомашнивания Aloes aleee ь. (Artiodactyla) на Костромской лесоферме. - "Первый межаунар. териолог, конгресс", М., 1974, с Джурович В.М., Михайлов А.П. Использование лосями древесной зелени. - "Охотничье хоз-о в интенс. компл. лесном хоз-е", Каунас-Гирионис, 1975, с. I20-I2I Дунин В.Ф. Химический состав и питательная ценность зимнего древесно-веточного корма лося. - В сб. "Березинский заповедник", вып. 3, Минск, "Ураджай", 1974, с И9. Дэвис Д.В., Христиан Дк.Ди. Популяционная регуляция у млекопитающих. - "Первый мехдунар. териол.конгресс", М., 1974, с Ельский Г.М. Типы популяции благородного оленя и косули в Украинских Карпатах. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с ирнов Д.В. Миграции лосей в Европейской части СССР. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, W 3, с Жирвов Д.В., Метельский А.П. О численности и промысле лосей в Московской области. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1965, 2, с Заблоцкая Д.В. Причины гибели лосей в различных географических районах. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, 3, с Заблоцкая Л.В. Динамика местных популяций лося на примере Подмосковья. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, Mi, 1970, с Заринов Р.3., Знаменский В.А. Численность, размещение и перспективы лосиного поголовья в ТАССР. - В сб. "Природн. ресурсы Волжско-Камского края", М., "Наука", 1964, с Зыкова Л.Ю. Движение численности и некоторые сведения по экологии лося в Окском заповеднике. - В сб."биол. и промысел лося", М., 1964, I, с


87 127. Зыков К.Д., Сапетин Я.В. Математическая модель распределения населения лосей. - В сб. "Структура и функционально- биогеоценотическая роль животного населения суши", И., 1967, с Иванаускас Т.Д. Состояние фауны Литвы и ее реконструкция. - В сб. "Научная сессия по вопросам биологии и сельского хозяйства", Рига, окт г.",м., 1953, с Иванов Ф.В. Методы учета охотничьих зверей и птиц в системе охотничьих хозяйств. - "Тр. 9-го Междунар. конгресса биологов-охотоведов, 1969", М., 1970, с Ивантер Э.В. Популяционная экология мелких млекопитающих таежного ceiepo запада СССР. Л., "Наука", Канаков Е.С. О соотношении полов в популяции лося Калининской области. - В сб. "Совр. состояние и пути развития охотовед, науки в СССР", Киров, 1974, с Киселев Р.П. К вопросу о соотношении полов в естественных популяциях животных. - "Тр. Окского государственного заповедника", "Лесн. пром-сть", М., 1967, вып. 7, с Киселева Е.Г., Приклонский С.Г., Теплов В.П. Материалы по размещению запасов лося, его плодовитости и соотношению полов. - "Тр. Окского гос. зап.", Воронеж, 1965, вып Кнорре Е.П. Экология лося. - "Тр. Печоро-Илычского гос. зап." Сыктывкар, Коми книжное изд-во, 1959, вып. 7, с Кнорре Е.П. Итоги и перспективы одомашнивания лося. - "Тр. Печеро-Илычского гос. зап.", 1961, вып Кожухов М.В. Итоги 20-летней экспериментальной работы по одомашниванию лося в Печоро-йлычском заповеднике. - В сб. "Одомашнивание лося", М., "Наука" 1973, с Козло П.Г. Динамика численности и морфологические особенности популяции Alcee alcee L. (Artiodactyla) Березинского заповедника. - "Первый ыеждунар. териолог. конгресс", М., 1974, с Козловский A.A. Лесные охотничьи угодья. М., "Лесн. пром-сть", 1971, с. I-I58.


88 139. Корнеев Г.А. Возможный механизм регуляции пола у грызунов. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, а., 1970,с Корочкина Д.Н. Факторы, влияющие на соотношение пола у зубров. - В сб. "Беловекск. пуща", Минск,"Ураджай", 1968, вып. 2, с Крайнев Е.Д. Некоторые итоги учета численности добываемой дичи на Украине. - Первая научная конф. по развитию охотн. хозяйства Укр. ССР, тезисы докладов Т, Киев, 1968, с Кубанцев Б.С. Условия существования и пол у млекопитающих. - "Уч. зап. Волгоградск. гос. пед. ин-та", 1964, вып. 16, с Кубанцев Б.С., Дьяконов В.Н., Рыбакова Т.И. Плотность популяции и динамика полового состава у млекопитающих. * "Экология", 1970, К; 3, с Лакин Г.ф. Биометрия. М.. "Высшая школа", 1968, с Ларин Б.А. О понятии "популяция" в прикладной экологии млекопитающих. '- В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., Г970, с. 8-И, Ларин Б.А. Концепции,популяционной структуры вида и охотничье хозяйство. - В сб. "Совр. состояние и пути развития охотовед, науки в СССР", Киров, т974, с Линг Х.И. Фауна промысловых млекопитающих ЗССР и пути ее реконструкции. - Автореф. дисс., Тарту, Линг Х.И. Изменения.численности и распространения лосей в Эстонии за последние столетия. - В ci. "География населения наземных животных и методы его изучения". Изд-во АН СССР, М., 1959, с Линг Х.И. Движение численности, распространение и плотность населения лосей в Эстонии. - В сб. "Биол. и промысел лося", -М., 1967, V 3, с Линг Х.И. Динамика про-дуктивности лося р Эстонии. -"Окология", J973, If 4, с I5T. Линг Х.И..Многоплодие, самок у лосей и продуктивность понуляпии. - Билл. МОИ 11, отд. яиил. 79(4),, с


89 152. Линг Х.И. Динамика структуры и продуктивности популяции Alcee alcee в Эстонии. - "Первый междунар. териол. конгресс", М., 1974, с Динг Х.И. Возможности использования данных о структуре популяции копытных и ее динамики для рациональной эксплуатации их запасов. - В сб. "Охотничье хоз-во в интенс. компл. лесном хоз-ве", Каунас-Гирионис, 1975, с Лихачев Г.Н. Лоси в Тульских засеках в I935-I95I гг. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1965, 2, с Дозан М.Н. Лоси в Молдавии. - "Природа", 1969, 12, с Лошкарев Г.А. Биологические группировки зайца-русака на Северном Кавказе. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с Майр Э. Зоологический вид и эволюция. М., "Мир", 1968, с Майр Э. Популяции, виды и эволюция. М., "Мир", 1974, с Макарова O.A. Некоторые данные по размножению лосей в Завидовском охотхозяйстве под Москвой. - "Уч. зап. Калининск. гос. пед. ин-та", т. 67, Х Малиновская Г.М. Химический состав листьев основных' древесно-веточных кормов, поедаемых оленем.- В сб. "Беловеж. пуща", Минск, "Ураджай", 1973, вып. 7, с Мартыненко H.A. Эмбриональная смертность сельскохозяйственных животных и ее предупреждение. Киев, "Урожай", 1971, с Михайлов А. Химический состав лесных кормов и их использование лосями. - МОИП, доклады 1970 и I полугодие 1971 г., 1972, с Михайлов А. Использование лосями летних кормов. -"Охота и охотн. х-во", 1972, 6, с Митинг Д. Охота в Эстонии. Таллин. 1965, с Насимович A.A. Роль режима снежного покрова в жизни копытных животных на территории СССР. М., Изд. Акад. наук СССР, 1955, с

.")
90 166. Насимович A.A. Используемая лосем территория. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1965, * 2, с Наумов Н.П. Структура популяции и динамика численности наземных позвоночных. - "Зоол. журнал", 1967, вып. 10, с Наумов H.H. Развитие представлений о популяциях в экологии животных. - В сб. "Очерки по истории экологии", М., "Наука", 1970, с Наумов Н.П. Пространственные структуры вида млекопитающих. - "Зоол. ж.", 1971, вып. 7, с Наумов Н.П. Теоретические основы и принципы экологии. - В сб. "Совр. пробл. экологии", М., 1973, с Наумов Н.П. Популяционная экология. Проблемы и задачи.- В сб. "Совр. пробл. экологии", М., 1973, с Наумов Н.П. Механизмы взаимодействия популяций (на примере некоторых млекопитающих и птиц). - "Зоол. ж.", 1975, вып. 6, с Наумов С.П., Гибет Л.А., Шаталова С.П. Динамика полового состава при изменениях численности млекопитающих. - "I. общ. биол.", 1969, 30, К" 6, с Новиков Г.А. Сто лет экологии Эрнста Геккеля. - В сб. "Очерки по истории экологии", М., "Наука", 1970, с Новиков Г.А., Айрапетьянц А.Э., Пукинский Ю.Б., Стрелков П.П., Тимофеева Е.К. Звери Ленинградской области. Л., Изд. Ленинградского ун-та, 1970, с Новиков Г.А., Иванов П.Д. Появление лосей в пределах Ленинграда. - "Бюлл. Моск. о-ва испыт. природы. Отд. биол.", 1970, т. 75, 6, с Паавер К.Л. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту, 1965, с Павлов М.П., Язан Ю.П. Случай многоплодия у лосей. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, 3, с Перовский М.Д. Потенциальная плодовитость лосей в различных ландшафтах. - В сб. "Совр. состояние и пути развития охотовед, науки в СССР", Киров, 1974, с


91 180. Петровский й.т. и др. Лоси в Витебской области Белоруссии. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, 3, с Петрусевич К. Об экологической организации популяции. - "Первый междунар. териол. конгресс", М., 1974, с Плохинский H.A. Биометрия. М., Изд. Моск. у-та, 1970, с Приклонский С.Г., Червонный В.В. Продуктивность лосиного стада в Европейской части РСФСР и возможности ее увеличения. - В сб. "Естественная производительность и продуктивность охотничьих угодий СССР",Мат. Всес. науч.-произведет, конф., часть I. Киров,1969, с Раков Н.В. Географические и экологические популяции лося в Амуро-Уссурийском крае. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1964, I, с Раков Н.В. Материалы по распространению и экологии лося в Амуро-Уссурийском крае. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1965, 2, с Раков Н.В. О значении некоторых факторов изоляции в проявлении и сохранении подвидовых признаков у лосей в Амуро-Уссурийском крае. - В сб.: Внутривидовая изменчивость наземных позвоночных животных и микроэволюция. Труды всесоюзного совещания, Свердловск, 1965, с Рафес U.M. Развитие учения о биогеоценозах. - В сб. "Очерки по истории экологии", У., "Наука", 1970, с Реймарс Н.Ф. Экологические сукцесси и промысловые животные. - В сб. "Охотоведение", М., "Лосн. пром-ть", с Розен Р. Принцип оптимальности в биологии. ГЛ., "Мир", 1969', с. I Русаков О.С. Двииение численности и распределение лося в Ленинградской области. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, 3, с


92 191. Русаков P.C. Распространение, ресурсы и хозяйственное использование копытных в северо-западных областях Европейской части СССР. - В сб. "Вопросы повышения продуктивности охотничьих угодий", М., "Колос", 1969, с Русаков P.C. Миграция лосей в северо-западных областях европейской части СССР. - Сб. науч.-тенх. информ. ВНИИ охот, х-ва и звероводства, 1973, т , с Смирнов B.C. Методы учета численности млекопитающих. - Тр. Ин-та биологии АН СССР Уральск, фил., вып. 39, Средне-Уральское книжное изд-во, 1964, с Смирнов B.C. Математические предпосылки учета численности охотничьих животных. - "Тр. Окского гос.зап.", вып. 9, 1973, с Смирнов М.Н. Территориальные группировки косули западного Забайкалья. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с Снедекор Дж. У. Статичтические методы в применении к исследованиям в сельском хозяйстве и биологии. М., Изд. сельскохоз. лит., жур. и плакатов, 1961, с. I Сысоев Н.Д. К биологической характеристике и перспективам использования популяции лося Владимировской области. - "Исслед. по фауне Сов. Союза", М., Теплов В.П. О принципах гибели лосей и факторах, влияющих на ее интенсивность. - "Зоол. ж.", т. 27, 1948, вып. I Теплов В.П. Динамика численности и годовые изменения в экологии промысловых животных Печорской тайги.-"тр. Печоро-Илычского гос. заповедника", вып. 8, Сыктывкар, Коми книжное изд-во, I960, с. I Теплова E.H. О стадности лосей в центральных областях РСФСР. - "Тр. Окского гос. зап.", вып. 7, И., "Лесн. пром-сть", 1967, с. II6-I Тимофеева Е.К. К методике изучения зимней экологии лося. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, R 3, С Тимофеева Е.К. Лось. Л., Изд. Ленингр. ун-та, 1974, с. I-I66. 93


93 203. Тимофеев-Ресовский Н.В., Яблоков A.B., Глотов Н.В.Очерк учения о популяции. И., "Наука", 1973, с Уатт К. Экология и управление природными ресурсами. М., "Мир", 1971, с Уильямсон М. Анализ биологических популяций. М., "Мир", 1975, с. 1-27I Федоров В.Д. Особенности организации биологических систем и гипотеза "вспышки" вида в сообществе. - Вестник Моск. ун-та, М., 1970, вып. 2, с Филонов К.П. Некоторые особенности естественной смертности копытных в заповедниках европейской части РСФСР. - В сб. "Охотничье хоз-о в интенс. компл. лесном хоз-е", Каунас-Гирионис, 1975, с Флинт В.Е. Типы пространственной структуры популяций у млекопитающих. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, У., 1970, с Флинт В.Е. О пространственных границах популяции и ее элементов у мелких млекопитающих. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с Флинт В.Е. Зональные особенности пространственной структуры популяции у мелких млекопитающих. - В сб."популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с Херувимов В.Д. Лось. Воронеж, Центрально-Черноземное книжное издательство, Червонный В.В. Об экологии, лесохозяйственном значении и промысле лося на Карельском перешейке. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1967, * 3, с Червонный В.В. Динамика численности лося на территорм Европейской части РСФСР. - Моск. общ. исп. природы. Доклады за П полугодие 1968 г. и 1969 г., М., 1971, с Шварц С.С. Принципы и методы современной экологии животных. - Тр. Йн-та биологии УФАН СССР, I960. вып Шварц С.С. Популяционная структура вица. - "Зоол. ж.", 1967, вып. 10, Cr


94 216. Шварц С.С. Эволюционная экология животных. - Тр. Ин-та экологии растений и животных АН СССР, Свердловск, 1969, с. I Шварц С.С. К истории основных понятий современной экологии. - В сб. "Очерки по истории экологии", М., "Наука", 1970, с Шварц С.С. Эколого-популяциоиные основы ведения охотничьего хозяйства. - "Тр. 9-го Междунар. конгресса биологов-охотоведов, 1969", М., 1970, с Шварц С.С. Актуальные проблемы учения о популяции и популяционная структура вида. - В сб. "Популяционная структура вида у млекопитающих", Моск. о-во исп. природы, М., 1970, с Шварц С.С. Теоретические основы и принципы экологии. - В сб. "Совр. пробл. экологии", М., 1973, с Шварц С.С. Эволюционная экология. - В сб. "Совр. пробл. экологии", М., 1973, с Шварц С.С. Биологические основы охотничьего хозяйства. - В сб. "Совр. состояние и пути развития охотовед, науки в СССР", Киров, 1974, с. 9-И Шубин Г.Г., Язан Ю.П. Опыт организации и ведения лосе- -промыслового хозяйства. - "Тр. Печоро-Илычского гос. зап.", Сыктывкар, Коми книжное изд-во, 1959, вып. 7, с Юргенсон П.Б. Роль фактора беспокойства в экологии зверей и птиц. - "Зоол. ж.", 1962, вып. 7, с Юргенсон П.Б. Структура и состав популяции лося в лесных охотничьих угодьях. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1964, I, с Юргенсон П.Б. Возрастная структура популяций и динамика плодовитости охотничьих животных. - "Зоол. ж.", 1966, вып. 2, с Юргенсон П.Б. Охотничьи звери и птицы. М., "Лесн. промыш-сть", 1968, с Юргенсон П.Б. Изменение в фауне охотничье-промысловых животных северного Подмосковья за последние лет. - Бюлл. МОИП, отд. биол., 1968, вып. 2, с


95 229. Юргенсон П.Б. Учет зимней численности охотничьих зверей и птиц по экскрементам. - "Тр. 9-го Междунар. конгресса биологов-охотоведов, 1969", М., 1970, с Юргенсон П.Б. Теория размещения, пространственный анализ и прикладная экология животных. - Бюлл. МОИП, отд. биол., 1970, вып. 6, с Юргенсон П.Б. Прикладное значение учения о популяциях охотничьих животных. - В сб. "Охотоведение", XXX, М., "Лесн. пром-сть", 1972, с Юргенсон П.Б. Биологические основы охотничьего хозяйства в лесах. М., "Лесн. пром-сть", 1973, с. I Юрлов К.Т. Об изменении ареала лося в пжной части Западно-Сибирской низменности. - В сб. "Биол» и промысел лося", М., Россельхозиздат, 1965, 2, с Язан Ю.П. Биологические особенности и пути хозяйственного освоения популяции мигрирующих лосей Печорской тайги. - "Тр. Печоро-Илычского гос. зап.", 196I, вып. 9, с. II4-20I Язан Ю.П. Плотности населения и показатели плодовитости лося Печорской таиги. - В сб. "Биол. и промысел лося", М., Россельхозиздат, 1964, I, с Язан Ю.П. Охотничьи звери печорской тайги (биология популяций, механизмы регуляции численности). Киров, Книгоиздат, 1972, с Язан Ю.П., Глушков В.М. Поведение лосей в снежный период. - Сб. научн.-техн. информ. ВНИИ охот, х-ва и звероводства, 1973, т , с Язан Ю.П., Кнорре Е.П. Проблема хозяйственного использования лосей в СССР. -"Тр. Печоро-Илычского гос. зап.", вып. II, Сыктывкар, Коми книжное изд-во, 1964, с Язан Ю.П., Кожухов М.В. Некоторые особенности терморегуляции у лосей. - "Тр. Печоро-Илычск. гос. зап.", 1964, вып. II, с
 так как структура популяции и происходящие в ней \"жизненные процессы\" по своей природе являются статистическими")

96 Дооь в Эстояи- Опыт системного анализа допуляц X. Лиг резюме При углубленном анвлеве етруктуры популяции лося исходили из следующего: 1) так как структура популяции и происходящие в ней "жизненные процессы" по своей природе являются статистическими явлениями, то необхцджм статистический анализ данных и использование методов, позволяющих получить для такого анализа пригодные материалы; 2) структура популяция и происходящие в ней "жизненные процессы" очень динамичны в пространстве и во времени и находятся под влиянием многих переменных; для выяснения действительных границ изменчивости и взаимосвязей необходим сбор данных sa длительный период; 3) так как популяция представляет собой сложную биологическую систему, то необходим системный подход, т.е. выяснение количественного состава субсистем и характер связей между ними в их динамике, в пространстве и во времени. Для анализа половой, возрастной, генерационной, социальной и территориальной структуры популяции лося использовали данные ЭстонииГ гг. о регистрации числа, пола и возраста (теленок или старше) лосей на осенних облавных охотах. За этот период собраны сведения об облавах в кварталах, где учтено групп и особей лося. Все данные проанализированы по пяти районам анализа, по годам и по трем месяцам. Вместе анализировали только данные, которые, используя критерий на достоверном уровне не различались. X2 использовали и при сравнении распределений между собой и с распределением Щгассона. Для оценки связей и силы корреляции использовали коэффициент корреляции г и rq. При выяснении системы размножения и продуктивности популяции лося использовались 10 показателей. Анализ динамик и 13 97


97 связей их с конкретными условиями позволил выявить ряд типов динамики показателей размножения. Последние позволяют прогнозировать продуктивность популяции, на основе которой рассматриваемый промежуток времени разделяется на три естественных периода: два из них (с 1961 по 1963 г. и 1968 по 1971г.) характеризуются высокой продуктивностью и третий (с 1964 по 1967 гг.) - низкой. В третьем периоде уровень продуктивности достиг уровня первого периода, но в то же время было меньше двоен и больше самок с телятами. Причиной этого было или омоложение популяции (вследствие охоты и иммиграции) или качественное различие генерации. Анализ продуктивности популяции из разных частей ареала показал, что потенциальная продуктивность одинакова на всем протяжении ареала, величина же ее реализации зависит от конкретных жизненных условий и структуры популяции. Сравнение собственных и литературных данных о наличии у лосих троен и четверен позволило утверждать, что это не случайное,а вполне закономерное явление. Частота их встречаемости не соответствует распределению Пуассона и является индикатором условий жизни и возрастной структуры популяции. Для выяснения переходного коэффициента между показателем плотности (особей на квартал) и абсолютной численности популяции сравнивали показатели плотности с официальными учетными данными, величиной популяции, которая вычислена на основании влияния отстрела на изменение показателя плотности и изменения возрастной структуры. На основе объективных данных реконструированная динамика численности заметно отличается от официальных данных: уже в 1970 году последние были на 5000 особей меньше действительного. Абсолютная плотность (особей на 1000 га) в 5,3 раза больше показателя плотности. Анализ динамики всех показателей размножения, продуктивности, плотности и динамики распространения и численности позволил заключить, что Эстонская популяция лося - открытая популяция, на которую заметное влияние оказывают иммиграция и эмиграция и обширные внутрипопуляционные перемещения. Для характеристики социальной структуры использовали данные о составе групп от I до 5 особей. Анализировали частоту встречаемости разных композиций по годам и месяцам. Выясненные закономерности позволяют данные социальной структуры исполь 98
\" группами наличие других социальных группировок не позволяет считать лося типично одиночным или оенейноживущим.")

98 зовать в качестве индикаторов продуктивности и возрастной структуры популяции. Наряду с одиночками и типичными "молочными (или материнскими)" группами наличие других социальных группировок не позволяет считать лося типично одиночным или оенейноживущим. Из территориальной структуры популяции анализировали распространение, встречаемость и динамику во времени и в пространстве группы особей, т.е. элементарные территориальные единицы популяции. Сравнение распространения разной величины групп в разные месяцы показало наличие двух типов распределения - осеннюю и зимнюю. Переход от одного типа к другому происходит в разные годы и в разные месяцы с разной интенсивностью. Анализ осеннего распределения показал, что на основе данных всей республики встречаемость групп разной величины (их распределение) не зависит от плотности и,следовательно, средняя величина групп в это время не может быть индикатором численности. За Il-летний период, по данным анализа пяти районов, осенью встречались 5 типов распределения, достоверно отличающиеся между собой. Два из них можно считать основными, один - переходный и два типа распределения возникли как результат внутрипопуляционных перемещений и поэтому можно использовать их в качестве индикаторов. Основными факторами,определяющими распределение,являются климатические условия; плотность влияет только тогда, когда лосем занята вся пригодная для его территория; значительно влияют на распределение внутрипопуляционные перемещения и изменения в социальной структуре; сравнение с Пуассоновским распределением показало их несовпадение на достоверном уровне во все месяцы, годы,а также и по районам анализа. Учитывая собранные в Эстонии данные о динамике размножения и структуре популяции лося, лишь на этом основании нельзя различатьвнутрипопуляционные группировки. На территории Эстонии нет основания подразделять животных на несколько популяций или внутрипопуляционных единиц (географических, локальных или элементарных популяций). Для характеристики структуры и продуктивности популяции часто используют слишком генерализированные показатели, которые искажают действительность и не позволяют выяснить структуру, продуктивность и дижамику их определяющих факто 99
99 ров. При детальном анализе структуры популяцшш я продуктивности незаменимы такие показатели-индикаторы, как плотность, фреквенто, распределение, доля самок с двойням! от всех самок и т.д. С точки зрения экологического аспекта на гомеостаз популяции надо смотреть не как на способность регуляции численности (плотности), а как на способность популяции поддерживать в равновесии свою структуру и противостоять факторам, пытающимся его изменить. Соответственно различаются три состояния гомеостаза: нормальный, экспансивный и зачищающий. Для рационального использования популяции лося в Эстонии необходимо уменьшить ее численность до лесохозяйстзенно допустимого уровня и, применяя правильные нормы отстрела и структуру отстрела, поддерживать популяцию в состоянии экспансивного гомеостаза. 100
 as the population structure and the processes going on within it are statistical phenomena in")

100 Ям Structure and Dynamics of the Population of Mow in Estonian SSB. An Attempt at a Svate^* AnnivaiB of the Population H. Ling summary The present deeper study into the population structure has proceeded fro* the following: 1) as the population structure and the processes going on within it are statistical phenomena in their essence, it is necessary to analyse the data statistically and to apply method» which make it possible to get materials corresponding to the requirements of a statistical analysist 2) the population structure and the processes in it are very dynamic both in tins and space and as they are Influenced by a lot of changing factors, it is indispensable to gather data on the actual limits and interrelations of the changes within the population as a whole and within its single eleaanta during a longer period of time; 2) in order to get acquainted with the population which is a complicated biological system, one must approach and analyse it systemically i.e. the quantitative composition of the sub-systems of the structure, the character and strength of their interrelations both in temporal and spatial changes have to be cleared up. For analysing the breeding of the moose population, its sex, age, generation, social and territorial structures, data were used that were collected during the autumn and winter battues of on the number of individuals, their sex and age (either a calf or older). Over a period of 11 years data were obtained on 53,96* chased 101


101 forest section of which 14,190 groups of noose totalling 43,566 individuals were driven out. All data on the five analysis districts for a threemonth period were annually analysed. Only the data that were not reliably different fron one another, were taken under a Joint observation. To ascertain the differences and similarities of the data belonging to different years, analysis districts and different months the Chi-square analysis was applied as a rule. When distributions were mutually compared or compared with the Poiseon distribution the same test was resorted to. For finding the relations and their strength the correlation coefficient r and rank correlation coefficient r 8 were used. In a number of cases the correlation matrices were calculated by the electronic computer "Minsk-32". Ten different indices were used to get information about the breeding and productivity of the moose population. The dynamics of these indices, the analysis of their interrelations and their association with concrete natural conditions made it possible to establish several types of breeding index dynamics. These indices can be applied when predicting the productivity of the population. On the basis of the latter three periods were distinguished: two periods of high fertility and a period of low fertility between them. Burning the third period nearly the equal level of productivity with that of the first period was reached. At the same time, however, the significance of females having twins had decreased while the importance of breeding females and those having only one calf had increased. The reasons may be either a change in the age structure of females (hunting,immigration) or the greater role played among the breeding females constitution owing to previous unfavourable conditions. The study of the respective data in the entire range was the same, its realisation, however, depended on concrete conditions. In Estonia, for example, in the period under observation there were years when productivity reached both its maximum and minimum throughout the are ad. The frequency of triplets and quadruplets and the investigation of data given in literature made it possible to 102


102 conclude that their occurrence is neither an accidental nor a rare phenomenon (does not correspond to the Poisson distribution) but rather regular and enables to use the frequency of triplets and quadruplets as an index of population's living conditions and its age structure. the For finding connections between the density index and the absolute size of the population, density was compared with official census data, with the population size calculated on the basis of information received from hunts on the basis of the relative index and finally compared with the size of the population calculated so as to take into account the change in the population* a age structure. The dynamics of population abundance reconstructed on the ground of objective data differed from official census data considerably: already in 1970 official census data were by about 5000 individuals smaller than the actual number of moose. The transition coefficient connecting the density index and population density was 5*3- i.e. population density was 5*3 times bigger than the density index. Considering all these indices and the dynamics of both distribution sind abundance it could be asserted that the Estonian moose population is a so-called open population whose structure and numerical dynamics were influenced in the studied period by both immigration and emigration as well as an extensive migration within the population. To get to know the social structure of the moose population and analysis of compositions of 1-5 individual groups was made on the basis of frequency of the following three components: old bulls, old cows and calves. The frequencies of occurrence and dynamics of different compositions per years were brought out. The same can be said about the regularities of frequencies of different compositions. These regularities can be used as indices of the age structure as well as the productivity of the population. Besides typical mother-families in normal moose population there exist constantly and in definite numerical relations other social groups. Therefore it is impossible to accept the hitherto valid standpoint as if the moose were a typical species living alone or only in mother families. 103


103 Of the population's territorial structure only the occurrence of co-existing and co-active groups (i.e. primary units using the territory) and the regularities of the dynamics of their temporal and spatial distributions were taken under investigation. The comparison of distributions of different months showed that there existed two distribution types: the autumn and the winter type. Transition fro» one type to the other takes place with different intensity in different years and at different times. The study of the autuim distribution type revealed that on the basis of data including the whole territory the frequency of moose herds of various sizes in October did not depend on the population density of moose. Consequently the average eise of the group cannot be the index of the population abundance. October distributions in analysis districts are most similar in West and North Estonia on the one hand, in Central, Bast and South Estonia on the other hand. Cobparing the October data of the whole observation period in analysis districts it turned out that during that period five essentially different distribution types occurred. Their temporal and spatial distributions showed definite regularities: two of them can be considered basic distribution types, one is a transition type and two types have formed as a result of migrations within the population. Consequently they can be taken as indices of the migrations. Comparison with neighbouring areas made it clear that in the distribution of moose herds typical of the Leningrad Begion was in good agreement with that of East Estonia (Alutaguse). The most significant factors determining the distribution are climatic conditions; the abundance of the population (population density) influences the distribution only when a certain density limit has been surpassed (after populating the areas of favourable living conditions); migrations within the population (the analysis of distributions enables to clear up their occurrence and direction) and social structure have some remarkable effect on the distribution; comparison with the Poieson distribution brought out reliable dia- 104

 on the territory")
104 tributional differences among aoose herds 1л all analysis districts, months and years. Taking into account the data collected on the dynamics of breeding and structure of the moose population, it is impossible to regard the differences either in productivity or population structures as the basis for differentiating the units within the population as it has been hitherto done. The analysis of our material shows that there are no grounds to differentiate several moose populations or subunits within the population (geographic,local or elementary populations) on the territory of Estonia. Too generalised assessments are often used as indices of both the population structure and population productivity. They distort reality and do not enable to find connections between different indices. Seither do they help to explain the structure, productivity or the factors directing their dynamics. Such indices as density, frequency, distribution, the role of females with twins among females having only one calf, etc. are indispensable in a detailed analysis of the population structure ae well as productivity. From the ecological aspect the homeostasis of the population must not be looked upon as the maintenance of the abundance level of the population but as the maintenance of the population structure. According to the states of the population also three states of homeostasis have been distinguished i.e. normal, expansive and protective states. In order to use the population rationally it is necessary to reduce the number of moose to the level permitted by our forest economy and to keep it in the state of expansive homeostasis by hunting the moose in proper numerical, age and sex proportions


105 kutitdd pötbadi taids к lim e stbdktddb a. talve PÖdra vanuse määramisel kasutati kahte meetodit. Ohe aluseks on hanroaste kulumisel paijanduva dentiiniriba laiuse ja kuju muutused looma vananemisel. Teine põhineb hambajuure mikrostruktuurl uurimisel, kus Jälgitakse tsemendi kihilist ladestumist. Talvekuudel (detsembrist märtsini) ladestunud kiht on tunduvalt tugevamini kaltsineerunud ja tihedam võrreldes ülejäänud ajal moodustunuga /4/. Kuna vaatluse all on hambajuure mikrostruktuur, tuleb sellest teha mikrolõigud, need värvida ja mikroskopeerida. Mikrolõigud võib teha alal8ualuu esimesest lõikehambast /2/ või esimesest piud. hambast /3/» Põdra puhul on puri hammast parem kasutada, sest tsemendi kiht selle hamba juurel on paksem ja pealegi saab ära kasutada mõlemad juuretipud, tehes Ühest piki, teisest ristlõigu."sel juhul oli parim määrand, smaterjal selline, kus peale lõikehammaste olid alles jäetud 2-3, purihammast. Meetod võimaldab vanust määrata aastase täpsusega kuni aastastel põtradel. Sellest east vanematel hakkavad esimesed tsemendiringid juba kustuma. Seda täpset, kuld töömahukat meetodit kasutati kontrollimaks, kas meil sobib kasutada taigavööndis põdra hammaste kulumise alusel koostatud vanuse määramise tabelit. Neil kahel meetodil тшratud materjali osas oli kokkulangevus küllalt suur (tabel 1). Saadud tulemused näitavad, et taiga-alal koostatud põdra vanuse määramise tabel /5/ on meil valdavalt kasutatav. Põdra vanuselist struktuuri uuriti 1962., 1970., ja aastal lastud loomade andmetel, kusjuures määrati valdavalt alalõualuu hammaste kulumisastme vanus järgi. Materjaliks oli põdra alalõualuu või selle osa, mis koguti TRÜ Zooloogiamuuseumi ja Eesti Metsainstituudi poolt. Sõltudes vanuse määramise viisist, tuli kogu materjal rühmita- 106

106 ,5-5, >8,5 Vanusrühmad Joon, 1. Vanuselise struktuuri muutused lastud põtrade andmetel aastatel 1962 ja

 Ф > TJ Saadud ваша tulemus 6 5 Saadud erinev tulemus с vx) ст\ i - «. vo vd <м 04 см m ф. 4» ö m 4» 3 ш Д cö cm о r- cm > -p vt) c 4 - c^> m ct\ ct\ ö\ сг\ as <")
107 .О eß 4 6N *- О СМ Т- 1 1А ОХ 1/4 со 9,5 - ' 43 ев w а VD o> <A VJD О CM CM IA r> ЧО IA T <" CM <Л t- IA IA o h- CM t- г- <т\ <* ^ cj см си g 6 а м о м 4,5-5,5 4,5-5,5 1А 5,5-1 см 1 и 3 4» О >а со см сг\ см о\ f- in т ftl г 00 сл г\ 40 см ч#- a\ frсо чр 4t см см см 8 тз р о ф и 2,5 2,5 LA la О 3,5 LA CM la 1 - г- 1 «- 1 LA О Ф ö g 4» Ю а +» о аз ьз со ЧО 4D см 1 см см см см о 40 л*- < со о г- о см гл i~ vo 4t ^ 1л СМ г- 1- СМ ^ О UD 1- ш 1 Ю гч fl Л Ih 2 0 9tS ^ (6 1 H 4» Л И ЛЗ н 'Н ф a Ф «н 4* 60 ш я? 9 f-a Я а) Ф > TJ Saadud ваша tulemus 6 5 Saadud erinev tulemus с vx) ст\ i - «. vo vd <м 04 см m ф. 4» ö m 4» 3 ш Д cö cm о r- cm > -p vt) c 4 - c^> m ct\ ct\ ö\ сг\ as <


108 de seile meetodi võimalustele vastavalt vanusegruppidesee (tabel 2). Kahe viimase aasta andmetes liideti 3,5-аавtaste rühm Järgmisega, sest ainult lõikehammaste järgi vanust määrates on nende kahe rühma vahel raske vahet teha. Vasikaid ja mullikaid (0,5- ja 1,5-aastased) a. ei laetud ning üle kümne aasta vanuseid loomi oli lastud nii vähe, et see rühm liideti eelmisega aasta kuulub põdrajahi algusaastatesse, kus põtru loendati nelja tuhande ringis ja valikut Jahil tehti hoolikalt, mida näitab seegi, et laetud loomade hulgas peaaegu puudusid mullikad, kuna laskmiseks valiti suured täiskasvanud loomad. Hoorte rohkus (2,5- ja 3,5-aastased) laetud isendite hulgas on seletatav nende suhteliselt suurema osaga looguslikus populatsioonis, kuna eelnevatelgi aastatel oli vasikate laskmine keelatud Ja mullikaid lasti üksikuid. Võrreldes aasta andmeid aastat hilisematega, ilmnevad lastud loomade vanuselises struktuuris suured muudatused. Olulised nihked on toimunud kõigis vanusrühmades (joon. 1). Hoorte osatähtsus on vähenenud, vanemate rühmade osa suurenenud. Suured erinevused üksikute vanusrühmade omavahelistes populatsioonides on kadunud. Kuna ja a. koondandmed statistiliselt erine (tabel 3), samuti mitte vanemad aastakäigud ei (aiatee 3,5-aastaetest) eraldi võetuna, siis võib nende andmeid koos vaadelda. Hoorte osas iseloomustab mõlemat aastat laetud vasikate ja mullikate väike protsent. Vasikate laskmine sõltub nende laskmiseks antud lubade arvust, mullikaid aga lastakse võimalikult vähem sellepärast, neid täiskasvanud loomadest ei eraldatud ning et laskestruktuuris eelistatakse lasta suuremat looma. Kõige vanema rühma küttimisel osa on suurenenud peamiselt üle kümne aasta vanuste arvel. Võrreldes ja aastate andmeid (alates 2,5 eluaastast) aastaga, on statistiline erinevus suur (X^2 = «40,25, df = 3, P < 0,05 %). Muutused, nagu juba eespool nimetatud, on toimunud struktuuri ühtlustumise suunas aastaga võrreldes on usaldatav erinevus koondandmetel (^2 = = 19,29, df = 6, P < 0,5 %), mis aga ilmselt on tingitud erinevusest kõige nooremates (0,5... 1,5-aastased) rühmades, sest selle osa väljajätmisel analüüsist ülejäänud osa struktuur oluliselt ei erine (tabel 3)


109 н * 4d «и 1 1 о 3 3 S н «> 3 н 'О о в k с 4* 4» «о о & tj g 9 й -Р -н н к 4» 15 x 4» в* 4* О сб «6 н 6 S-I ti H Ж О «А о о с\ СМ РЧ л л л 1 M * <л с\ «л ГЛ 00 чо xh 00 <л см <D <ф чо (Л u> ta * О О о РЧ О О О о О т V V л А Л я H r\ «л яг M * ta ft ft ft ft й гл IA t- О 4h GO CM о см см в м IA õ t- чо h- d *< CM <d tit > <н h td IA IA IA о см «л СМ О л V v ч/ ia чо чо чо # #» ft из 1А <0 оч 04 *sh О см ia 1А ih C7N К <7\ <Т\ XJ eö + + > CM о см СМ СМ о 03 г- D- с- t- е- с- tj cn o> сг> Огч ОХ СГ\ H XJ (0 4» CM CM о О см fi to <0 t- Г- с~~ г~ <o cd (Tv (Jx СП СГх (TN ст. > ti T ~ T т-


110 1972. aaata kohta laekunud materjal jaotub vanusrühaadeeee kõige proportsionaalsemalt. Hinnatav on vasikate laskenormi suurenemine. Mullikaid laeti euhtelieelt vähem, kui neid esineb populatsioonis. On Jätkunud kõige vanema rühma osatähtsuse tõus, mis oli täheldatav Juba a. alates. Esitatud andmetest võib teha järgmised järeldused: 1. Lastud loomade vanuseline struktuur ei kajasta otseselt looduses esineva populatsiooni vanuselist struktuuri, kuna esineb valikuline küttimine. Valiku põhjuste Ja valiku suuna arvestamine võimaldab aga ka laetud loomade vanuse määramisel saadud andmeid kasutada loodusliku populatsiooni vanuselise struktuuri indikaatorina. 2. Kütitud põtrade hulgas on aastatel võrreldes aastaga märksa suurenenud vanemate loomade osa, kuid seni kogutud andmed ei võimalda väita, et samasuunaliselt on muutunud ka loodusliku populatsiooni struktuur. 3. Kütitud põtrade vanuselise struktuuri selgitamine võimaldab kontrollida kasutatava laskenormi ja -struktuuri Õigsust. Tuleb pidada hädavajalikuks kütitavate ulukite (põder, metssiga, metskits) alalõugade-hammaste kogumist, nende iga-aastast läbitöötamist ja tulemuste kasutamist laskenormi ja -struktuuri määramisel. 4. Käesoleva ajani põtrade küttimisel rakendatav lubade süsteem ei taga populatsiooni õige vanuselise struktuuri säilitamist. K a s u t a t u d k i r j a n d u s 1. L a w s R. M. A g e d e t e r m i n a t i o n o f p i n n i p e d e s w i t h special reference to growth layers in the teeth. - "Säugetiere", 1962, Bd. 27, Hr. 3, S L o w W. A. A g e d e t e r m i n a t i o n o f d e e r b y a n n u l a r s t r u c ture of dental cementum. - Dep. of Zoology, the Univ. of Brit. Columbia, Vancouver,
111 3. M i t c h e l l В. D e t e r m i n a t i o n o f a g e i n e c o t t i e h red deer from growth layers in dental cement. - "Nature", 1963, 198, p К л е в е з а л ь Т. А., К л е й н б е р г C. E. О п р е деление возраста млекопитающих по слоистым структурам зубов и кости. - М., "Наука", 1967, с. I-I К н о р р е Е. П., Ш у б и н Т. Г. О п р е д е л е н и е в о з раста лося. - "Тр. Лечоро-Илычского гос. зап.", Сыктывкар, Коми книжное изд., вып. 7, 1959, с


112 Возрастная структура отстрелянных лосей А. Талве р е з ю м е Для определения возраста лосей по стёртости зубов на 15 экземплярах сравнивали картину стёртости с микроструктурой корней зубов. По степени стёртости зубов определили возраст у 591 особи. Сравнивая данные с 1970 по 1972 год с 1962 годом, заметно увеличение старших возрастных групп.данные 1971 и 1972 года достоверно не различаются.хотя возрастная струитура отстрелянных лосей отличается от таковой хе живой популяции по причине выборочного отстрела, эти данные позволяют проверить правильность используемой нормы и структуры отстрела. The Age Structure of the Shot Moose A. Talve s u m m a r y To determine the age of the moose by the degree of wear of his teeth the accordance of wear with the age ascertained through the analysis of the micro structure of the tooth root was checked in 15 specimens. Thus the age of 591 moose was found out. Comparing the material with that of 1962 it became clear that the significance of older animals was considerably greater in The data of 1970 and 1971 showed no reliable discrepancy. A1thought the age structure of the shot animals differs from that in a natural population owing to selective hunting. These data make it possible to verify the correctness of the numerical, age and sex proportions of shooting. 113

113 SISUKORD. СОДЕРЖАНИЕ. CONTENTS H. Ling. PSdrapopulatsloonl struktuur ja dünaamika Beatl HSV-a. Populatsiooni süsteemse analüüsi kat ee 3 X. Линг. Структура и динамика популяции лося в Эстонской ССР. Опыт системного анализа популяц и и. Р е з ю м е 9 7 Н. Ling. The Structure end Dynamics of the Population of Moose in Estonian SSR. An Attempt at a Systemic Analysis of the Population. Summar у 101 A. Talve. Kutitud põtrade vanuseline struktuur А. Талве. Возрастная структура отстрелянных лосей. Ре эюм е A. Talve. The Age Structure of the Shot Moose. из S u m m г у 113
Microsoft Word - Järvamaa_KOVid_rahvastiku analüüs.doc
 Töömaterjal. Rivo Noorkõiv. Käesolev töö on koostatud Siseministeeriumi poolt osutatava kohalikeomavalitsuste ühinemist toetava konsultatsioonitöö raames. Järvamaa omavalitsuste rahvastiku arengu üldtrendid
Töömaterjal. Rivo Noorkõiv. Käesolev töö on koostatud Siseministeeriumi poolt osutatava kohalikeomavalitsuste ühinemist toetava konsultatsioonitöö raames. Järvamaa omavalitsuste rahvastiku arengu üldtrendid
raamat5_2013.pdf
 Peatükk 5 Prognoosiintervall ja Usaldusintervall 5.1 Prognoosiintervall Unustame hetkeks populatsiooni parameetrite hindamise ja pöördume tagasi üksikvaatluste juurde. On raske ennustada, milline on huvipakkuva
Peatükk 5 Prognoosiintervall ja Usaldusintervall 5.1 Prognoosiintervall Unustame hetkeks populatsiooni parameetrite hindamise ja pöördume tagasi üksikvaatluste juurde. On raske ennustada, milline on huvipakkuva
my_lauluema
 Lauluema Lehiste toomisel A. Annisti tekst rahvaluule õhjal Ester Mägi (1983) Soran Alt q = 144 Oh se da ke na ke va de ta, ae ga i lust üü ri kes ta! üü ri kes ta! 3 Ju ba on leh tis lei na kas ke, hal
Lauluema Lehiste toomisel A. Annisti tekst rahvaluule õhjal Ester Mägi (1983) Soran Alt q = 144 Oh se da ke na ke va de ta, ae ga i lust üü ri kes ta! üü ri kes ta! 3 Ju ba on leh tis lei na kas ke, hal
Microsoft Word - Uudiskirja_Toimetulekutoetus docx
 Toimetulekutoetuse maksmine 2014. 2018. aastal Sotsiaalministeeriumi analüüsi ja statistika osakond Toimetulekutoetust on õigus saada üksi elaval isikul või perekonnal, kelle kuu netosissetulek pärast
Toimetulekutoetuse maksmine 2014. 2018. aastal Sotsiaalministeeriumi analüüsi ja statistika osakond Toimetulekutoetust on õigus saada üksi elaval isikul või perekonnal, kelle kuu netosissetulek pärast
Matemaatiline analüüs IV 1 3. Mitme muutuja funktsioonide diferentseerimine 1. Mitme muutuja funktsiooni osatuletised Üleminekul ühe muutuja funktsioo
 Matemaatiline analüüs IV 1 3. Mitme muutuja funktsioonide diferentseerimine 1. Mitme muutuja funktsiooni osatuletised Üleminekul üe muutuja funktsioonidelt m muutuja funktsioonidele, kus m, 3,..., kerkib
Matemaatiline analüüs IV 1 3. Mitme muutuja funktsioonide diferentseerimine 1. Mitme muutuja funktsiooni osatuletised Üleminekul üe muutuja funktsioonidelt m muutuja funktsioonidele, kus m, 3,..., kerkib
(Microsoft Word - S\365raliste seisund Eestis 2009.doc)
") Keskkonnaministeerium Keskkonnateabe Keskus Uluksõraliste asurkondade seisund Eestis 2009 KIK 2009. a jahindusprogrammi projekti nr 1 lõpparuanne sõraliste seire spetsialist: Jüri Tõnisson teadustöö spetsialist:
Keskkonnaministeerium Keskkonnateabe Keskus Uluksõraliste asurkondade seisund Eestis 2009 KIK 2009. a jahindusprogrammi projekti nr 1 lõpparuanne sõraliste seire spetsialist: Jüri Tõnisson teadustöö spetsialist:
Excel Valemite koostamine (HARJUTUS 3) Selles peatükis vaatame millistest osadest koosnevad valemid ning kuidas panna need Excelis kirja nii, et
 Selles peatükis vaatame millistest osadest koosnevad valemid ning kuidas panna need Excelis kirja nii, et") Excel2016 - Valemite koostamine (HARJUTUS 3) Selles peatükis vaatame millistest osadest koosnevad valemid ning kuidas panna need Excelis kirja nii, et programm suudaks anda tulemusi. Mõisted VALEM - s.o
Excel2016 - Valemite koostamine (HARJUTUS 3) Selles peatükis vaatame millistest osadest koosnevad valemid ning kuidas panna need Excelis kirja nii, et programm suudaks anda tulemusi. Mõisted VALEM - s.o
Antennide vastastikune takistus
 Antennide vastastikune takistus Eelmises peatükis leidsime antenni kiirgustakistuse arvestamata antenni lähedal teisi objekte. Teised objektid, näiteks teised antennielemendid, võivad aga mõjutada antenni
Antennide vastastikune takistus Eelmises peatükis leidsime antenni kiirgustakistuse arvestamata antenni lähedal teisi objekte. Teised objektid, näiteks teised antennielemendid, võivad aga mõjutada antenni
Matemaatilised meetodid loodusteadustes. I Kontrolltöö I järeltöö I variant 1. On antud neli vektorit: a = (2; 1; 0), b = ( 2; 1; 2), c = (1; 0; 2), d
, b = ( 2; 1; 2), c = (1; 0; 2), d") Matemaatilised meetodid loodusteadustes I Kontrolltöö I järeltöö I variant On antud neli vektorit: a (; ; ), b ( ; ; ), c (; ; ), d (; ; ) Leida vektorite a ja b vaheline nurk α ning vekoritele a, b ja
Matemaatilised meetodid loodusteadustes I Kontrolltöö I järeltöö I variant On antud neli vektorit: a (; ; ), b ( ; ; ), c (; ; ), d (; ; ) Leida vektorite a ja b vaheline nurk α ning vekoritele a, b ja
Institutsioonide usaldusväärsuse uuring
 INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet I kvartal 01 Liis Grünberg Pärnu mnt, 1 Tallinn +() 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet I kvartal 01 Liis Grünberg Pärnu mnt, 1 Tallinn +() 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
2016 aasta märtsi tulumaksu laekumine omavalitsustele See ei olnud ette arvatav Tõesti ei olnud, seda pole juhtunud juba tükk aega. Graafikult näeme,
 2016 märtsi tulumaksu laekumine omavalitsustele See ei olnud ette arvatav Tõesti ei olnud, seda pole juhtunud juba tükk aega. Graafikult näeme, et märtsis laekus tulumaksu eelmise märtsist vähem ka 2009
2016 märtsi tulumaksu laekumine omavalitsustele See ei olnud ette arvatav Tõesti ei olnud, seda pole juhtunud juba tükk aega. Graafikult näeme, et märtsis laekus tulumaksu eelmise märtsist vähem ka 2009
Tallinna Tehnikakõrgkooli teadustegevuse kajastus Eesti Teadusinfosüsteemi andmebaasis Agu Eensaar, füüsika-matemaatikakandidaat Eesti Teadusinfosüste
 Tallinna Tehnikakõrgkooli teadustegevuse kajastus Eesti Teadusinfosüsteemi andmebaasis Agu Eensaar, füüsika-matemaatikakandidaat Eesti Teadusinfosüsteem (ETIS) (https://www.etis.ee/index.aspx) kajastab
Tallinna Tehnikakõrgkooli teadustegevuse kajastus Eesti Teadusinfosüsteemi andmebaasis Agu Eensaar, füüsika-matemaatikakandidaat Eesti Teadusinfosüsteem (ETIS) (https://www.etis.ee/index.aspx) kajastab
VL1_praks6_2010k
 Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is oma kursuse ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht (Insert / Lisa -> Worksheet / Tööleht), nimetage
Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is oma kursuse ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht (Insert / Lisa -> Worksheet / Tööleht), nimetage
Suunised Euroopa turu infrastruktuuri määruse (EMIR) kohaste kesksetele vastaspooltele suunatud protsüklilisusvastaste tagatismeetmete kohta 15/04/201
 kohaste kesksetele vastaspooltele suunatud protsüklilisusvastaste tagatismeetmete kohta 15/04/201") Suunised Euroopa turu infrastruktuuri määruse (EMIR) kohaste kesksetele vastaspooltele suunatud protsüklilisusvastaste tagatismeetmete kohta 15/04/2019 ESMA70-151-1496 ET Sisukord I. Reguleerimisala...
Suunised Euroopa turu infrastruktuuri määruse (EMIR) kohaste kesksetele vastaspooltele suunatud protsüklilisusvastaste tagatismeetmete kohta 15/04/2019 ESMA70-151-1496 ET Sisukord I. Reguleerimisala...
Euroopa Liidu tulevik aastal 2013 Euroopa Liidu tulevikust räägitakse kõikjal ja palju, on tekkinud palju küsimusi ning levib igasugust valeinfot, mis
 Euroopa Liidu tulevik aastal 2013 Euroopa Liidu tulevikust räägitakse kõikjal ja palju, on tekkinud palju küsimusi ning levib igasugust valeinfot, mis ajab inimesed segadusse. Järgnevalt on ülevaade mõningatest
Euroopa Liidu tulevik aastal 2013 Euroopa Liidu tulevikust räägitakse kõikjal ja palju, on tekkinud palju küsimusi ning levib igasugust valeinfot, mis ajab inimesed segadusse. Järgnevalt on ülevaade mõningatest
Institutsioonide usaldusväärsuse uuring
 INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet I kvartal 0 Liis Grünberg Pärnu mnt, Tallinn +() 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet I kvartal 0 Liis Grünberg Pärnu mnt, Tallinn +() 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
Polünoomi juured Juure definitsioon ja Bézout teoreem Vaadelgem polünoomi kus K on mingi korpus. f = a 0 x n + a 1 x n a n 1 x
 1 5.5. Polünoomi juured 5.5.1. Juure definitsioon ja Bézout teoreem Vaadelgem polünoomi kus K on mingi korpus. f = a 0 x n + a 1 x n 1 +... + a n 1 x + a n K[x], (1) Definitsioon 1. Olgu c K. Polünoomi
1 5.5. Polünoomi juured 5.5.1. Juure definitsioon ja Bézout teoreem Vaadelgem polünoomi kus K on mingi korpus. f = a 0 x n + a 1 x n 1 +... + a n 1 x + a n K[x], (1) Definitsioon 1. Olgu c K. Polünoomi
(Microsoft Word - Purgatsi j\344rve supluskoha suplusvee profiil l\374hike)
") PURGATSI JÄRVE SUPLUSKOHA SUPLUSVEE PROFIIL Harjumaa, Aegviidu vald Koostatud: 01.03.2011 Täiendatud 19.09.2014 Järgmine ülevaatamine: vastavalt vajadusele või veekvaliteedi halvenemisel 1 Purgatsi järve
PURGATSI JÄRVE SUPLUSKOHA SUPLUSVEE PROFIIL Harjumaa, Aegviidu vald Koostatud: 01.03.2011 Täiendatud 19.09.2014 Järgmine ülevaatamine: vastavalt vajadusele või veekvaliteedi halvenemisel 1 Purgatsi järve
VKE definitsioon
 Väike- ja keskmise suurusega ettevõtete (VKE) definitsioon vastavalt Euroopa Komisjoni määruse 364/2004/EÜ Lisa 1-le. 1. Esiteks tuleb välja selgitada, kas tegemist on ettevõttega. Kõige pealt on VKE-na
Väike- ja keskmise suurusega ettevõtete (VKE) definitsioon vastavalt Euroopa Komisjoni määruse 364/2004/EÜ Lisa 1-le. 1. Esiteks tuleb välja selgitada, kas tegemist on ettevõttega. Kõige pealt on VKE-na
Automaatjuhtimise alused Automaatjuhtimissüsteemi kirjeldamine Loeng 2
 Automaatjuhtimise alused Automaatjuhtimissüsteemi kirjeldamine Loeng 2 Laplace'i teisendus Diferentsiaalvõrrandite lahendamine ilma tarkvara toeta on keeruline Üheks lahendamisvõtteks on Laplace'i teisendus
Automaatjuhtimise alused Automaatjuhtimissüsteemi kirjeldamine Loeng 2 Laplace'i teisendus Diferentsiaalvõrrandite lahendamine ilma tarkvara toeta on keeruline Üheks lahendamisvõtteks on Laplace'i teisendus
Tala dimensioonimine vildakpaindel
 Tala dimensioonimine vildakpaindel Ülesanne Joonisel 9 kujutatud okaspuidust konsool on koormatud vertikaaltasandis ühtlase lauskoormusega p ning varda teljega risti mõjuva kaldjõuga (-jõududega) F =pl.
Tala dimensioonimine vildakpaindel Ülesanne Joonisel 9 kujutatud okaspuidust konsool on koormatud vertikaaltasandis ühtlase lauskoormusega p ning varda teljega risti mõjuva kaldjõuga (-jõududega) F =pl.
EDL Liiga reeglid 1. ÜLDSÄTTED 1.1. EDL Liiga toimub individuaalse arvestuse alusel, kus mängijad on jagatud hooaja EDL Liiga tulemuste põhj
 EDL Liiga reeglid 1. ÜLDSÄTTED 1.1. EDL Liiga toimub individuaalse arvestuse alusel, kus mängijad on jagatud hooaja 2017-2018 EDL Liiga tulemuste põhjal nelja liigasse. a. Premium Liiga (9 osalejat) b.
EDL Liiga reeglid 1. ÜLDSÄTTED 1.1. EDL Liiga toimub individuaalse arvestuse alusel, kus mängijad on jagatud hooaja 2017-2018 EDL Liiga tulemuste põhjal nelja liigasse. a. Premium Liiga (9 osalejat) b.
FIE Jaanus Elts Metsakaitse- ja Metsauuenduskeskus Töövõtulepingu nr 2-24/Trt-17, 7. aprill 2008 aruanne Metskurvitsa mängulennu seire aastal Ja
 FIE Jaanus Elts Metsakaitse- ja Metsauuenduskeskus Töövõtulepingu nr 2-24/Trt-17, 7. aprill 2008 aruanne Metskurvitsa mängulennu seire 2008. aastal Jaanus Elts Tartu, 2008 Metskurvits on erakordselt raskesti
FIE Jaanus Elts Metsakaitse- ja Metsauuenduskeskus Töövõtulepingu nr 2-24/Trt-17, 7. aprill 2008 aruanne Metskurvitsa mängulennu seire 2008. aastal Jaanus Elts Tartu, 2008 Metskurvits on erakordselt raskesti
loeng7.key
 Grammatikate elustamine JFLAPiga Vesal Vojdani (TÜ Arvutiteaduse Instituut) Otse Elust: Java Spec https://docs.oracle.com/javase/specs/jls/se8/html/ jls-14.html#jls-14.9 Kodutöö (2. nädalat) 1. Avaldise
Grammatikate elustamine JFLAPiga Vesal Vojdani (TÜ Arvutiteaduse Instituut) Otse Elust: Java Spec https://docs.oracle.com/javase/specs/jls/se8/html/ jls-14.html#jls-14.9 Kodutöö (2. nädalat) 1. Avaldise
Mida me teame? Margus Niitsoo
 Mida me teame? Margus Niitsoo Tänased teemad Tagasisidest Õppimisest TÜ informaatika esmakursuslased Väljalangevusest Üle kogu Ülikooli TÜ informaatika + IT Kokkuvõte Tagasisidest NB! Tagasiside Tagasiside
Mida me teame? Margus Niitsoo Tänased teemad Tagasisidest Õppimisest TÜ informaatika esmakursuslased Väljalangevusest Üle kogu Ülikooli TÜ informaatika + IT Kokkuvõte Tagasisidest NB! Tagasiside Tagasiside
vv05lah.dvi
 IMO 05 Eesti võistkonna valikvõistlus 3. 4. aprill 005 Lahendused ja vastused Esimene päev 1. Vastus: π. Vaatleme esiteks juhtu, kus ringjooned c 1 ja c asuvad sirgest l samal pool (joonis 1). Olgu O 1
IMO 05 Eesti võistkonna valikvõistlus 3. 4. aprill 005 Lahendused ja vastused Esimene päev 1. Vastus: π. Vaatleme esiteks juhtu, kus ringjooned c 1 ja c asuvad sirgest l samal pool (joonis 1). Olgu O 1
Relatsiooniline andmebaaside teooria II. 6. Loeng
 Relatsiooniline andmebaaside teooria II. 5. Loeng Anne Villems ATI Loengu plaan Sõltuvuste pere Relatsiooni dekompositsioon Kadudeta ühendi omadus Sõltuvuste pere säilitamine Kui jõuame, siis ka normaalkujud
Relatsiooniline andmebaaside teooria II. 5. Loeng Anne Villems ATI Loengu plaan Sõltuvuste pere Relatsiooni dekompositsioon Kadudeta ühendi omadus Sõltuvuste pere säilitamine Kui jõuame, siis ka normaalkujud
Lisa I_Müra modelleerimine
 LISA I MÜRA MODELLEERIMINE Lähteandmed ja metoodika Lähteandmetena kasutatakse AS K-Projekt poolt koostatud võimalikke eskiislahendusi (trassivariandid A ja B) ning liiklusprognoosi aastaks 2025. Kuna
LISA I MÜRA MODELLEERIMINE Lähteandmed ja metoodika Lähteandmetena kasutatakse AS K-Projekt poolt koostatud võimalikke eskiislahendusi (trassivariandid A ja B) ning liiklusprognoosi aastaks 2025. Kuna
Microsoft Word - QOS_2008_Tallinn_OK.doc
 GSM mobiiltelefoniteenuse kvaliteet Tallinnas, juuni 2008 Sideteenuste osakond 2008 Kvaliteedist üldiselt GSM mobiiltelefonivõrgus saab mõõta kümneid erinevaid tehnilisi parameetreid ja nende kaudu võrku
GSM mobiiltelefoniteenuse kvaliteet Tallinnas, juuni 2008 Sideteenuste osakond 2008 Kvaliteedist üldiselt GSM mobiiltelefonivõrgus saab mõõta kümneid erinevaid tehnilisi parameetreid ja nende kaudu võrku
Praks 1
 Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is oma kursuse ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht, nimetage see ümber leheküljeks Praks6 ja 3.
Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is oma kursuse ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht, nimetage see ümber leheküljeks Praks6 ja 3.
EUROOPA KOMISJON Brüssel, COM(2018) 284 final ANNEXES 1 to 2 LISAD järgmise dokumendi juurde: Ettepanek: Euroopa Parlamendi ja nõukogu määru
 284 final ANNEXES 1 to 2 LISAD järgmise dokumendi juurde: Ettepanek: Euroopa Parlamendi ja nõukogu määru") EUROOPA KOMISJON Brüssel, 17.5.2018 COM(2018) 284 final ANNEXES 1 to 2 LISAD järgmise dokumendi juurde: Ettepanek: Euroopa Parlamendi ja nõukogu määrus, millega kehtestatakse uute raskeveokite CO2-heite
EUROOPA KOMISJON Brüssel, 17.5.2018 COM(2018) 284 final ANNEXES 1 to 2 LISAD järgmise dokumendi juurde: Ettepanek: Euroopa Parlamendi ja nõukogu määrus, millega kehtestatakse uute raskeveokite CO2-heite
Andmed arvuti mälus Bitid ja baidid
 Andmed arvuti mälus Bitid ja baidid A bit about bit Bitt, (ingl k bit) on info mõõtmise ühik, tuleb mõistest binary digit nö kahendarv kahe võimaliku väärtusega 0 ja 1. Saab näidata kahte võimalikku olekut
Andmed arvuti mälus Bitid ja baidid A bit about bit Bitt, (ingl k bit) on info mõõtmise ühik, tuleb mõistest binary digit nö kahendarv kahe võimaliku väärtusega 0 ja 1. Saab näidata kahte võimalikku olekut
lvk04lah.dvi
 Lahtine matemaatikaülesannete lahendamise võistlus. veebruaril 004. a. Lahendused ja vastused Noorem rühm 1. Vastus: a) jah; b) ei. Lahendus 1. a) Kuna (3m+k) 3 7m 3 +7m k+9mk +k 3 3M +k 3 ning 0 3 0,
Lahtine matemaatikaülesannete lahendamise võistlus. veebruaril 004. a. Lahendused ja vastused Noorem rühm 1. Vastus: a) jah; b) ei. Lahendus 1. a) Kuna (3m+k) 3 7m 3 +7m k+9mk +k 3 3M +k 3 ning 0 3 0,
AS TEEDE TEHNOKESKUS LIIKLUSLOENDUS LIIKLUSSAGEDUSKÕVERAD TUGIMAANTEEDEL Tallinn 2001
 AS TEEDE TEHNOKESKUS LIIKLUSLOENDUS LIIKLUSSAGEDUSKÕVERAD TUGIMAANTEEDEL Tallinn 21 LIIKLUSSAGEDUSKÕVERAD TUGIMAANTEEDEL Projektijuht: Kristjan Duubas AS Teede Tehnokeskus Leping 29.3.21 SISUKORD 1. Saateks
AS TEEDE TEHNOKESKUS LIIKLUSLOENDUS LIIKLUSSAGEDUSKÕVERAD TUGIMAANTEEDEL Tallinn 21 LIIKLUSSAGEDUSKÕVERAD TUGIMAANTEEDEL Projektijuht: Kristjan Duubas AS Teede Tehnokeskus Leping 29.3.21 SISUKORD 1. Saateks
Institutsioonide usaldusväärsuse uuring
 INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet II kvartal 01 Liis Grünberg Pärnu mnt, 1 Tallinn +() 55 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
INSTITUTSIOONIDE USALDUSVÄÄRSUS Maksu- ja Tolliamet II kvartal 01 Liis Grünberg Pärnu mnt, 1 Tallinn +() 55 0 Liis@turu-uuringute.ee www.turu-uuringute.ee METOODIKA Tulemuste omandiõigus: kuulub Turu-uuringuta
Praks 1
 Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht, nimetage see ümber leheküljeks Praks6 ja 3. kopeerige
Biomeetria praks 6 Illustreeritud (mittetäielik) tööjuhend Eeltöö 1. Avage MS Excel is ankeedivastuseid sisaldav andmestik, 2. lisage uus tööleht, nimetage see ümber leheküljeks Praks6 ja 3. kopeerige
Non-pharmacological treatment
 Siinusrütmi säilitava ravimi valik Kliiniline küsimus Kas siinusrütmi säilitavaks raviks tuleks eelistada mõnd konkreetset ravimirühma/ravimit: BBL vs Ic vs III? Olulised tulemusnäitajad Surm, ajuinfarkt,
Siinusrütmi säilitava ravimi valik Kliiniline küsimus Kas siinusrütmi säilitavaks raviks tuleks eelistada mõnd konkreetset ravimirühma/ravimit: BBL vs Ic vs III? Olulised tulemusnäitajad Surm, ajuinfarkt,
Markina
 EUROOPA NOORTE ALKOHOLITARBIMISE PREVENTSIOONI PRAKTIKAD JA SEKKUMISED Anna Markina Tartu Ülikool Meie ülesanne on: Tuvastada ja välja valida erinevaid programme ja sekkumist, mida on hinnatud ja mille
EUROOPA NOORTE ALKOHOLITARBIMISE PREVENTSIOONI PRAKTIKAD JA SEKKUMISED Anna Markina Tartu Ülikool Meie ülesanne on: Tuvastada ja välja valida erinevaid programme ja sekkumist, mida on hinnatud ja mille
(10. kl. I kursus, Teisendamine, kiirusega, kesk.kiirusega \374lesanded)
") TEISENDAMINE Koostanud: Janno Puks 1. Massiühikute teisendamine Eesmärk: vajalik osata teisendada tonne, kilogramme, gramme ja milligramme. Teisenda antud massiühikud etteantud ühikusse: a) 0,25 t = kg
TEISENDAMINE Koostanud: Janno Puks 1. Massiühikute teisendamine Eesmärk: vajalik osata teisendada tonne, kilogramme, gramme ja milligramme. Teisenda antud massiühikud etteantud ühikusse: a) 0,25 t = kg
Traneksaam_ortopeedias
 Traneksaamhape kasutamine Juri Karjagin Tartu Ülikool Tartu Ülikooli Kliinikum Plaan Fibrinolüüsist Traneksaamhape farmakoloogiast Traneksaamhape uuringud Plaaniline kirurgia Erakorraline trauma Toopiline
Traneksaamhape kasutamine Juri Karjagin Tartu Ülikool Tartu Ülikooli Kliinikum Plaan Fibrinolüüsist Traneksaamhape farmakoloogiast Traneksaamhape uuringud Plaaniline kirurgia Erakorraline trauma Toopiline
Microsoft PowerPoint - Loodusteaduslik uurimismeetod.ppt
 Bioloogia Loodusteaduslik uurimismeetod Tiina Kapten Bioloogia Teadus, mis uurib elu. bios - elu logos - teadmised Algselt võib rääkida kolmest teadusharust: Botaanika Teadus taimedest Zooloogia Teadus
Bioloogia Loodusteaduslik uurimismeetod Tiina Kapten Bioloogia Teadus, mis uurib elu. bios - elu logos - teadmised Algselt võib rääkida kolmest teadusharust: Botaanika Teadus taimedest Zooloogia Teadus
Tartu Ülikool
 Tartu Ülikool Code coverage Referaat Koostaja: Rando Mihkelsaar Tartu 2005 Sissejuhatus Inglise keelne väljend Code coverage tähendab eesti keeles otse tõlgituna koodi kaetust. Lahti seletatuna näitab
Tartu Ülikool Code coverage Referaat Koostaja: Rando Mihkelsaar Tartu 2005 Sissejuhatus Inglise keelne väljend Code coverage tähendab eesti keeles otse tõlgituna koodi kaetust. Lahti seletatuna näitab
Microsoft PowerPoint - Loeng2www.ppt [Compatibility Mode]
![Microsoft PowerPoint - Loeng2www.ppt [Compatibility Mode]](/thumbs/88/116353219.jpg "Microsoft PowerPoint - Loeng2www.ppt [Compatibility Mode]") Biomeetria 2. loeng Lihtne lineaarne regressioon mudeli hindamisest; usaldusintervall; prognoosiintervall; determinatsioonikordaja; Märt Möls martm@ut.ee Y X=x~ N(μ=10+x; σ=2) y 10 15 20 2 3 4 5 6 7 8
Biomeetria 2. loeng Lihtne lineaarne regressioon mudeli hindamisest; usaldusintervall; prognoosiintervall; determinatsioonikordaja; Märt Möls martm@ut.ee Y X=x~ N(μ=10+x; σ=2) y 10 15 20 2 3 4 5 6 7 8
Microsoft Word - Vorm_TSD_Lisa_1_juhend_2015
 TSD lisa 1 täitmise juhend Olulisemad muudatused deklareerimisel alates 01.01.2015 vorm TSD lisal 1. Alates 01.01.2015 muutus vorm TSD ja tema lisad. Deklaratsioonivorme muutmise peamine eesmärk oli tagada
TSD lisa 1 täitmise juhend Olulisemad muudatused deklareerimisel alates 01.01.2015 vorm TSD lisal 1. Alates 01.01.2015 muutus vorm TSD ja tema lisad. Deklaratsioonivorme muutmise peamine eesmärk oli tagada
Tartu Ülikool Matemaatika-informaatikateaduskond Matemaatilise statistika instituut Võrgupeo külastaja uurimine Andmeanalüüs I projekt Koostajad: Urma
 Tartu Ülikool Matemaatika-informaatikateaduskond Matemaatilise statistika instituut Võrgupeo külastaja uurimine Andmeanalüüs I projekt Koostajad: Urmas Kvell Riivo Talviste Gert Palok Juhendaja: Mare Vähi
Tartu Ülikool Matemaatika-informaatikateaduskond Matemaatilise statistika instituut Võrgupeo külastaja uurimine Andmeanalüüs I projekt Koostajad: Urmas Kvell Riivo Talviste Gert Palok Juhendaja: Mare Vähi
1
 IDA-TALLINNA KESKHAIGLA Statsionaarsete patsientide rahulolu uuring 13 13 Sisukord 1. Uuringu läbiviimise metoodika... 2 2. Andmete analüüs... 2 3. Uuringu valim... 3 3.1. Vastanute iseloomustus: sugu,
IDA-TALLINNA KESKHAIGLA Statsionaarsete patsientide rahulolu uuring 13 13 Sisukord 1. Uuringu läbiviimise metoodika... 2 2. Andmete analüüs... 2 3. Uuringu valim... 3 3.1. Vastanute iseloomustus: sugu,
Abiarstide tagasiside 2016 Küsimustikule vastas 137 tudengit, kellest 81 (60%) olid V kursuse ning 56 (40%) VI kursuse tudengid. Abiarstina olid vasta
 olid V kursuse ning 56 (40%) VI kursuse tudengid. Abiarstina olid vasta") Abiarstide tagasiside 2016 Küsimustikule vastas 137 tudengit, kellest 81 (60%) olid V kursuse ning 56 (40%) VI kursuse tudengid. Abiarstina olid vastanutest töötanud 87 tudengit ehk 64%, kellest 79 (91%)
Abiarstide tagasiside 2016 Küsimustikule vastas 137 tudengit, kellest 81 (60%) olid V kursuse ning 56 (40%) VI kursuse tudengid. Abiarstina olid vastanutest töötanud 87 tudengit ehk 64%, kellest 79 (91%)
Matemaatika ainekava 8.klass 4 tundi nädalas, kokku 140 tundi Kuu Õpitulemus Õppesisu Algebra (65 t.) Geomeetria (60 t.) Ajavaru kordamiseks (15 õppet
 Geomeetria (60 t.) Ajavaru kordamiseks (15 õppet") Matemaatika ainekava 8.klass 4 tundi nädalas, kokku 140 tundi Algebra (65 t.) Geomeetria (60 t.) Ajavaru kordamiseks (15 õppetundi) septembernovember korrastab hulkliikmeid Hulkliige. Tehted liidab, lahutab
Matemaatika ainekava 8.klass 4 tundi nädalas, kokku 140 tundi Algebra (65 t.) Geomeetria (60 t.) Ajavaru kordamiseks (15 õppetundi) septembernovember korrastab hulkliikmeid Hulkliige. Tehted liidab, lahutab
Solaariumisalongides UVseadmete kiiritustiheduse mõõtmine. Tallinn 2017
 Solaariumisalongides UVseadmete kiiritustiheduse mõõtmine. Tallinn 2017 1. Sissejuhatus Solaariumides antakse päevitusseansse kunstliku ultraviolettkiirgusseadme (UV-seadme) abil. Ultraviolettkiirgus on
Solaariumisalongides UVseadmete kiiritustiheduse mõõtmine. Tallinn 2017 1. Sissejuhatus Solaariumides antakse päevitusseansse kunstliku ultraviolettkiirgusseadme (UV-seadme) abil. Ultraviolettkiirgus on
Ruutvormid Denitsioon 1. P n Ütleme, et avaldis i;j=1 a ijx i x j ; kus a ij = a ji ; a ij 2 K ja K on korpus, on ruutvorm üle korpuse K muutujate x 1
 Ruutvormid Denitsioon. P n Ütleme, et avaldis i;j= a ijx i x j ; kus a ij = a ji ; a ij K ja K on korus, on ruutvorm üle koruse K muutujate x ;;x n suhtes. Maatriksit =(a ij ) nimetame selle ruutvormi
Ruutvormid Denitsioon. P n Ütleme, et avaldis i;j= a ijx i x j ; kus a ij = a ji ; a ij K ja K on korus, on ruutvorm üle koruse K muutujate x ;;x n suhtes. Maatriksit =(a ij ) nimetame selle ruutvormi
Microsoft PowerPoint - VKP_VÜFdial_J_AnnikaUettekanne_VKP_ _taiendatudMU.ppt [Compatibility Mode]
![Microsoft PowerPoint - VKP_VÜFdial_J_AnnikaUettekanne_VKP_ _taiendatudMU.ppt [Compatibility Mode]](/thumbs/98/137859934.jpg "Microsoft PowerPoint - VKP_VÜFdial_J_AnnikaUettekanne_VKP_ _taiendatudMU.ppt [Compatibility Mode]") Kuidas arendada kohalikke avalikke teenuseid omavalitsuste ja kodanikuühenduste koostöös? Annika Uudelepp Praxise juhatuse liige, Valitsemise ja kodanikeühiskonna programmi direktor 16.09.2009 Tallinnas
Kuidas arendada kohalikke avalikke teenuseid omavalitsuste ja kodanikuühenduste koostöös? Annika Uudelepp Praxise juhatuse liige, Valitsemise ja kodanikeühiskonna programmi direktor 16.09.2009 Tallinnas
Microsoft PowerPoint - Keskkonnamoju_rus.ppt
 Keskkonnakonverents 07.01.2011 Keskkonnamõju hindamine ja keskkonnamõju strateegiline hindamine on avalik protsess kuidas osaleda? Elar Põldvere (keskkonnaekspert, Alkranel OÜ) Kõik, mis me õpime täna,
Keskkonnakonverents 07.01.2011 Keskkonnamõju hindamine ja keskkonnamõju strateegiline hindamine on avalik protsess kuidas osaleda? Elar Põldvere (keskkonnaekspert, Alkranel OÜ) Kõik, mis me õpime täna,
(Tõrked ja töökindlus \(2\))
)") Elektriseadmete tõrked ja töökindlus Click to edit Master title style 2016 sügis 2 Prof. Tõnu Lehtla VII-403, tel.6203 700 http://www.ttu.ee/energeetikateaduskond/elektrotehnika-instituut/ Kursuse sisu
Elektriseadmete tõrked ja töökindlus Click to edit Master title style 2016 sügis 2 Prof. Tõnu Lehtla VII-403, tel.6203 700 http://www.ttu.ee/energeetikateaduskond/elektrotehnika-instituut/ Kursuse sisu
Lasteendokrinoloogia aktuaalsed küsimused
 Haigusjuht nooruki androloogiast lasteendokrinoloogi pilgu läbi Aleksandr Peet SA TÜK Lastekliinik aleksandr.peet@kliinikum.ee Neuroloogi jälgimisel vanuseni 13 a. Esmakordselt visiidil vanuses 7a Elu
Haigusjuht nooruki androloogiast lasteendokrinoloogi pilgu läbi Aleksandr Peet SA TÜK Lastekliinik aleksandr.peet@kliinikum.ee Neuroloogi jälgimisel vanuseni 13 a. Esmakordselt visiidil vanuses 7a Elu
Komisjoni delegeeritud määrus (EL) nr 862/2012, 4. juuni 2012, millega muudetakse määrust (EÜ) nr 809/2004 seoses teabega nõusoleku kohta prospekti ka
 nr 862/2012, 4. juuni 2012, millega muudetakse määrust (EÜ) nr 809/2004 seoses teabega nõusoleku kohta prospekti ka") L 256/4 Euroopa Liidu Teataja 22.9.2012 MÄÄRUSED KOMISJONI DELEGEERITUD MÄÄRUS (EL) nr 862/2012, 4. juuni 2012, millega muudetakse määrust (EÜ) nr 809/2004 seoses teabega nõusoleku kohta prospekti kasutamiseks,
L 256/4 Euroopa Liidu Teataja 22.9.2012 MÄÄRUSED KOMISJONI DELEGEERITUD MÄÄRUS (EL) nr 862/2012, 4. juuni 2012, millega muudetakse määrust (EÜ) nr 809/2004 seoses teabega nõusoleku kohta prospekti kasutamiseks,
PISA 2015 tagasiside koolile Tallinna Rahumäe Põhikool
 PISA 215 tagasiside ile Tallinna Rahumäe Põhi PISA 215 põhiuuringus osales ist 37 õpilast. Allpool on esitatud ülevaade i õpilaste testisoorituse tulemustest. Võrdluseks on ära toodud vastavad näitajad
PISA 215 tagasiside ile Tallinna Rahumäe Põhi PISA 215 põhiuuringus osales ist 37 õpilast. Allpool on esitatud ülevaade i õpilaste testisoorituse tulemustest. Võrdluseks on ära toodud vastavad näitajad
Saksa keele riigieksamit asendavate eksamite tulemuste lühianalüüs Ülevaade saksa keele riigieksamit asendavatest eksamitest Saksa keele riigi
 Saksa keele riigieksamit asendavate eksamite tulemuste lühianalüüs 2014 1. Ülevaade saksa keele riigieksamit asendavatest eksamitest Saksa keele riigieksam on alates 2014. a asendatud Goethe-Zertifikat
Saksa keele riigieksamit asendavate eksamite tulemuste lühianalüüs 2014 1. Ülevaade saksa keele riigieksamit asendavatest eksamitest Saksa keele riigieksam on alates 2014. a asendatud Goethe-Zertifikat
Lisa 7.1. KINNITATUD juhatuse a otsusega nr 2 MTÜ Saarte Kalandus hindamiskriteeriumite määratlemine ja kirjeldused 0 nõrk e puudulik -
 Lisa 7.1. KINNITATUD juhatuse 04. 01. 2018. a otsusega nr 2 MTÜ Saarte Kalandus hindamiskriteeriumite määratlemine ja kirjeldused 0 nõrk e puudulik - kriteerium ei ole täidetud (hindepunkti 0 saab rakendada
Lisa 7.1. KINNITATUD juhatuse 04. 01. 2018. a otsusega nr 2 MTÜ Saarte Kalandus hindamiskriteeriumite määratlemine ja kirjeldused 0 nõrk e puudulik - kriteerium ei ole täidetud (hindepunkti 0 saab rakendada
Microsoft Word - Manage PODER 2011 RJfin.doc
 Keskkonnateabe Keskus Ulukiseireosakond Eesti põdraasurkonna seisundist ja uurimisest Küttimisettepanek 2011. aastaks Jüri Tõnisson Rauno Veeroja Tartu 2011 Sisukord Saateks... 2 Eesti põdraasurkonna seisundist
Keskkonnateabe Keskus Ulukiseireosakond Eesti põdraasurkonna seisundist ja uurimisest Küttimisettepanek 2011. aastaks Jüri Tõnisson Rauno Veeroja Tartu 2011 Sisukord Saateks... 2 Eesti põdraasurkonna seisundist
Põhja-Harju Koostöökogu HINDAMISKRITEERIUMID Kinnitatud üldkoosoleku otsusega p 2.2. Hindamiskriteeriumid I III MEEDE Osakaal % Hinne Selgi
 Hindamiskriteeriumid I III MEEDE Osakaal % Hinne Selgitus Viide projektikirjeldusele Projekti ettevalmistuse ja elluviimise kvaliteediga seotud kriteeriumid (kokku 0%) 1. Projekti sidusus ja põhjendatus
Hindamiskriteeriumid I III MEEDE Osakaal % Hinne Selgitus Viide projektikirjeldusele Projekti ettevalmistuse ja elluviimise kvaliteediga seotud kriteeriumid (kokku 0%) 1. Projekti sidusus ja põhjendatus
II lisa Ravimi omaduste kokkuvõtte ja pakendi infolehe muudatused, esitatud Euroopa Ravimiameti poolt Käesolev ravimi omaduste kokkuvõte ja pakendi in
 II lisa Ravimi omaduste kokkuvõtte ja pakendi infolehe muudatused, esitatud Euroopa Ravimiameti poolt Käesolev ravimi omaduste kokkuvõte ja pakendi infoleht on esildismenetluse tulemus. Vastavalt vajadusele
II lisa Ravimi omaduste kokkuvõtte ja pakendi infolehe muudatused, esitatud Euroopa Ravimiameti poolt Käesolev ravimi omaduste kokkuvõte ja pakendi infoleht on esildismenetluse tulemus. Vastavalt vajadusele
Õppekava arendus
 Õppekava arendus Ülle Liiber Õppekava kui kokkulepe ja ajastu peegeldus Riiklik õppekava on peegeldus sellest ajast, milles see on koostatud ja kirjutatud valitsevast mõtteviisist ja inimkäsitusest, pedagoogilistest
Õppekava arendus Ülle Liiber Õppekava kui kokkulepe ja ajastu peegeldus Riiklik õppekava on peegeldus sellest ajast, milles see on koostatud ja kirjutatud valitsevast mõtteviisist ja inimkäsitusest, pedagoogilistest
untitled
 IDA-VIRUMAA PÕLEVKIVI TÖÖSTUSSE SUHTUMISE UURINGU ARUANNE IDA-VIRUMAA ELANIKKONNA TELEFONIKÜSITLUS Oktoober 2006 www.saarpoll.ee SISUKORD 1. Sissejuhatus ja metoodika........... 3 2. Põhijäreldused....
IDA-VIRUMAA PÕLEVKIVI TÖÖSTUSSE SUHTUMISE UURINGU ARUANNE IDA-VIRUMAA ELANIKKONNA TELEFONIKÜSITLUS Oktoober 2006 www.saarpoll.ee SISUKORD 1. Sissejuhatus ja metoodika........... 3 2. Põhijäreldused....
2015 aasta veebruari tulumaksu laekumise lühianalüüs aasta veebruari lühianalüüs pole eriti objektiivne, sest veebruari lõpuks polnud tuludeklar
 2015 aasta i tulumaksu laekumise lühianalüüs. 2015 aasta i lühianalüüs pole eriti objektiivne, sest i lõpuks polnud tuludeklaratsioonid laekunud veel üle 2500 ettevõttelt. Rahandusministeerium püüdis küll
2015 aasta i tulumaksu laekumise lühianalüüs. 2015 aasta i lühianalüüs pole eriti objektiivne, sest i lõpuks polnud tuludeklaratsioonid laekunud veel üle 2500 ettevõttelt. Rahandusministeerium püüdis küll
PowerPoint Presentation
 LEOSTUMINE Transpiratsioon Leostumine Evaporatsioon Eestis on sademete hulk aastas umbes 1,5 korda aurumisest suurem. Keskmiselt on meil sademeid 550-800 mm ja aurub 320-440 mm aastas (. Maastik) Seniste
LEOSTUMINE Transpiratsioon Leostumine Evaporatsioon Eestis on sademete hulk aastas umbes 1,5 korda aurumisest suurem. Keskmiselt on meil sademeid 550-800 mm ja aurub 320-440 mm aastas (. Maastik) Seniste
Microsoft Word - L_5_2018_docx.docx
 Maaeluministri 0.0.07 määrus nr 4 Põllumajandusettevõtja tulemuslikkuse parandamise investeeringutoetus Lisa (maaeluministri. novembri 08 määruse nr 6 sõnastuses) Teravilja, õliseemnete ja valgurikaste
Maaeluministri 0.0.07 määrus nr 4 Põllumajandusettevõtja tulemuslikkuse parandamise investeeringutoetus Lisa (maaeluministri. novembri 08 määruse nr 6 sõnastuses) Teravilja, õliseemnete ja valgurikaste
Microsoft Word - essee_CVE ___KASVANDIK_MARKKO.docx
 Tartu Ülikool CVE-2013-7040 Referaat aines Andmeturve Autor: Markko Kasvandik Juhendaja : Meelis Roos Tartu 2015 1.CVE 2013 7040 olemus. CVE 2013 7040 sisu seisneb krüptograafilises nõrkuses. Turvaaugu
Tartu Ülikool CVE-2013-7040 Referaat aines Andmeturve Autor: Markko Kasvandik Juhendaja : Meelis Roos Tartu 2015 1.CVE 2013 7040 olemus. CVE 2013 7040 sisu seisneb krüptograafilises nõrkuses. Turvaaugu
Sügis 2018 Kõrgema matemaatika 2. kontrolltöö tagasiside Üle 20 punkti kogus tervelt viis üliõpilast: Robert Johannes Sarap, Enely Ernits, August Luur
 Sügis 2018 Kõrgema matemaatika 2. kontrolltöö tagasiside Üle 20 punkti kogus tervelt viis üliõpilast: Robert Johannes Sarap, Enely Ernits, August Luure, Urmi Tari ja Miriam Nurm. Ka teistel oli edasiminek
Sügis 2018 Kõrgema matemaatika 2. kontrolltöö tagasiside Üle 20 punkti kogus tervelt viis üliõpilast: Robert Johannes Sarap, Enely Ernits, August Luure, Urmi Tari ja Miriam Nurm. Ka teistel oli edasiminek
Monitooring 2010f
 Lõimumiskava monitooring 2010 Raivo Vetik, TLÜ võrdleva poliitika professor Kohtumine Rahvuste Ümarlauas 24. september, 2010 Uuringu taust TLÜ uurimisgrupp: Raivo Vetik, Jüri Kruusvall, Maaris Raudsepp,
Lõimumiskava monitooring 2010 Raivo Vetik, TLÜ võrdleva poliitika professor Kohtumine Rahvuste Ümarlauas 24. september, 2010 Uuringu taust TLÜ uurimisgrupp: Raivo Vetik, Jüri Kruusvall, Maaris Raudsepp,
Lisa 2 Maanteeameti peadirektori käskkirjale nr 0250 Kattega riigimaanteede taastusremondi objektide valikumetoodika Maanteeamet Tallinn 20
 Lisa 2 Maanteeameti peadirektori 02.07.2013 käskkirjale nr 0250 Kattega riigimaanteede taastusremondi objektide valikumetoodika Maanteeamet Tallinn 2013 0 Sisukord Sisukord... 1 Sissejuhatus... 2 Metoodika
Lisa 2 Maanteeameti peadirektori 02.07.2013 käskkirjale nr 0250 Kattega riigimaanteede taastusremondi objektide valikumetoodika Maanteeamet Tallinn 2013 0 Sisukord Sisukord... 1 Sissejuhatus... 2 Metoodika
Renovation of Historic Wooden Apartment Buildings
 Hoonete õhuleke ja selle mõõtmine Click to edit Master title style Endrik Arumägi Targo Kalamees Teadmistepõhine ehitus 26.04.2018 Piirdetarindite õhulekked Iseloomustavad suurused õhuvahetuvuskordsus
Hoonete õhuleke ja selle mõõtmine Click to edit Master title style Endrik Arumägi Targo Kalamees Teadmistepõhine ehitus 26.04.2018 Piirdetarindite õhulekked Iseloomustavad suurused õhuvahetuvuskordsus
IMO 2000 Eesti võistkonna valikvõistlus Tartus, aprillil a. Ülesannete lahendused Esimene päev 1. Olgu vaadeldavad arvud a 1, a 2, a 3,
 IMO 000 Eesti võistkonna valikvõistlus Tartus, 19. 0. aprillil 000. a. Ülesannete lahendused Esimene päev 1. Olgu vaadeldavad arvud a 1, a, a 3, a 4, a 5. Paneme tähele, et (a 1 + a + a 3 a 4 a 5 ) (a
IMO 000 Eesti võistkonna valikvõistlus Tartus, 19. 0. aprillil 000. a. Ülesannete lahendused Esimene päev 1. Olgu vaadeldavad arvud a 1, a, a 3, a 4, a 5. Paneme tähele, et (a 1 + a + a 3 a 4 a 5 ) (a
Microsoft Word - Errata_Andmebaaside_projekteerimine_2013_06
 Andmebaaside projekteerimine Erki Eessaar Esimene trükk Teadaolevate vigade nimekiri seisuga 24. juuni 2013 Lehekülg 37 (viimane lõik, teine lause). Korrektne lause on järgnev. Üheks tänapäeva infosüsteemide
Andmebaaside projekteerimine Erki Eessaar Esimene trükk Teadaolevate vigade nimekiri seisuga 24. juuni 2013 Lehekülg 37 (viimane lõik, teine lause). Korrektne lause on järgnev. Üheks tänapäeva infosüsteemide
Microsoft PowerPoint - Mart2.ppt
 Üldindeksi järjestuse tutvustamine ja põhjendamine Mart Uba Biomeetria sektor mart.uba@jkkeskus.ee Üldindeksi arvutamine Sobivaks üldindeksi mudeliks kõrge elupäevatoodanguga lehmade eluajatoodangu suurendamiseks
Üldindeksi järjestuse tutvustamine ja põhjendamine Mart Uba Biomeetria sektor mart.uba@jkkeskus.ee Üldindeksi arvutamine Sobivaks üldindeksi mudeliks kõrge elupäevatoodanguga lehmade eluajatoodangu suurendamiseks
(Microsoft Word - \334levaade erakondade finantsseisust docx)
") Ülevaade erakondade finantsmajanduslikust olukorrast seisuga 31.12.2010 Ülevaate eesmärgiks on kirjeldada erakondade rahalist seisu, mis annab informatsiooni nende tugevusest või nõrkusest, mis omakorda
Ülevaade erakondade finantsmajanduslikust olukorrast seisuga 31.12.2010 Ülevaate eesmärgiks on kirjeldada erakondade rahalist seisu, mis annab informatsiooni nende tugevusest või nõrkusest, mis omakorda
Microsoft Word - 4. Haiguse mõõtmine.doc
 HAIGUSE ESINEMISSAGEDUSE NÄITAJAD "Epidemioloogia on (murru) nimetajate teadus ning sellena kasulik vastukaal kliinilisele orientatsioonile, mida peamiselt valitsevad (murru) lugejad. Nimetajate sissetoomine
HAIGUSE ESINEMISSAGEDUSE NÄITAJAD "Epidemioloogia on (murru) nimetajate teadus ning sellena kasulik vastukaal kliinilisele orientatsioonile, mida peamiselt valitsevad (murru) lugejad. Nimetajate sissetoomine
R4BP 3 Print out
 Biotsiidi omaduste kokkuvõte Biotsiidi nimi: Dismate PE Biotsiidi liik (liigid): Tooteliik 9 - Repellendid ja atraktandid (kahjuritõrje) Loa number: UK-06-08 Biotsiidiregistri (R4BP 3) kande viitenumber:
Biotsiidi omaduste kokkuvõte Biotsiidi nimi: Dismate PE Biotsiidi liik (liigid): Tooteliik 9 - Repellendid ja atraktandid (kahjuritõrje) Loa number: UK-06-08 Biotsiidiregistri (R4BP 3) kande viitenumber:
Microsoft Word - 03_ausus lisaylesanded.doc
 ÕPL LS 3 LSÜLSNDD USUS ML eemat usus (sh teisi teemasid) saab sisse juhatada ka HHK- (H HLB KSULK) meetodil. Näiteks: Miks on ausus hea? Miks on ausus halb? Miks on ausus kasulik? H: Hoiab ära segadused
ÕPL LS 3 LSÜLSNDD USUS ML eemat usus (sh teisi teemasid) saab sisse juhatada ka HHK- (H HLB KSULK) meetodil. Näiteks: Miks on ausus hea? Miks on ausus halb? Miks on ausus kasulik? H: Hoiab ära segadused
Word Pro - digiTUNDkaug.lwp
 / näide: \ neeldumisseadusest x w x y = x tuleneb, et neeldumine toimub ka näiteks avaldises x 2 w x 2 x 5 : x 2 w x 2 x 5 = ( x 2 ) w ( x 2 ) [ x 5 ] = x 2 Digitaalskeemide optimeerimine (lihtsustamine)
/ näide: \ neeldumisseadusest x w x y = x tuleneb, et neeldumine toimub ka näiteks avaldises x 2 w x 2 x 5 : x 2 w x 2 x 5 = ( x 2 ) w ( x 2 ) [ x 5 ] = x 2 Digitaalskeemide optimeerimine (lihtsustamine)
Microsoft Word - ref - Romet Piho - Tutorial D.doc
 Tartu Ülikool Andmetöötluskeel "Tutorial D" realisatsiooni "Rel" põhjal Referaat aines Tarkvaratehnika Romet Piho Informaatika 2 Juhendaja Indrek Sander Tartu 2005 Sissejuhatus Tänapäeval on niinimetatud
Tartu Ülikool Andmetöötluskeel "Tutorial D" realisatsiooni "Rel" põhjal Referaat aines Tarkvaratehnika Romet Piho Informaatika 2 Juhendaja Indrek Sander Tartu 2005 Sissejuhatus Tänapäeval on niinimetatud
Tööplaan 9. kl õpik
 Mõttest tekstini Eesti keele ja tekstiõpetuse õpik 9. klassile Näidistööplaan Aeg Teema Põhimõisted Õppematerjal Tegevused Õppetulemus Hindamine 1. nädal I. Suhtlemine rühmas Ptk 1 Sissejuhatuseks 2. nädal
Mõttest tekstini Eesti keele ja tekstiõpetuse õpik 9. klassile Näidistööplaan Aeg Teema Põhimõisted Õppematerjal Tegevused Õppetulemus Hindamine 1. nädal I. Suhtlemine rühmas Ptk 1 Sissejuhatuseks 2. nädal
ÕPETAJATE OSKUSED PIAAC ANDMETE BAASIL Aune Valk PIAAC (Programme for the International Assessment of Adult Competencies) uuringu raames va
 uuringu raames va") ÕPETAJATE OSKUSED PIAAC ANDMETE BAASIL 10.12.2013 Aune Valk PIAAC (Programme for the International Assessment of Adult Competencies) uuringu raames valmis väga rikas andmebaas, mis annab võimaluse uurida
ÕPETAJATE OSKUSED PIAAC ANDMETE BAASIL 10.12.2013 Aune Valk PIAAC (Programme for the International Assessment of Adult Competencies) uuringu raames valmis väga rikas andmebaas, mis annab võimaluse uurida
Suunised Reitinguagentuuride meetodite valideerimise ja läbivaatamise suunised 23/03/2017 ESMA/2016/1575 ET
 Suunised Reitinguagentuuride meetodite valideerimise ja läbivaatamise suunised 23/03/2017 ESMA/2016/1575 ET Sisukord 1 Kohaldamisala... 3 2 Mõisted, õiguslikud viited ja lühendid... 4 3 Eesmärk... 5 4
Suunised Reitinguagentuuride meetodite valideerimise ja läbivaatamise suunised 23/03/2017 ESMA/2016/1575 ET Sisukord 1 Kohaldamisala... 3 2 Mõisted, õiguslikud viited ja lühendid... 4 3 Eesmärk... 5 4
Individuaalne õppekava ja selle koostamine I. ÜLDSÄTTED Kehtestatud dir kk nr 32/ Individuaalse õppekava koostamise aluseks on 1.
 Individuaalne õppekava ja selle koostamine I. ÜLDSÄTTED Kehtestatud dir kk 20.11.2012 nr 32/1.1-6 1.1. Individuaalse õppekava koostamise aluseks on 1.1.1. Põhikooli ja gümnaasiumiseadus 18. Vastu võetud
Individuaalne õppekava ja selle koostamine I. ÜLDSÄTTED Kehtestatud dir kk 20.11.2012 nr 32/1.1-6 1.1. Individuaalse õppekava koostamise aluseks on 1.1.1. Põhikooli ja gümnaasiumiseadus 18. Vastu võetud
PowerPoint Presentation
 Strateegilise koostöö projekti eelarve Katriin Ranniku 17.02.2016 Millest tuleb juttu? Mis reguleerib Erasmus+ programmist rahastatavaid projekte? Millised on Erasmus+ strateegilise koostöö projekti eelarveread?
Strateegilise koostöö projekti eelarve Katriin Ranniku 17.02.2016 Millest tuleb juttu? Mis reguleerib Erasmus+ programmist rahastatavaid projekte? Millised on Erasmus+ strateegilise koostöö projekti eelarveread?
Võrguväljaanded ja veebiarhiveerimine
 e24.ee folklore.ee delfi.ee www.ut.ee www.delfi.ee ut.ee Teeme ISE: harilikud hoidised virtuaalsetest viljadest veebiarhiivi riiulil Jaanus Kõuts, Eesti Rahvusraamatukogu 17.09.2013 Kogemused http://archive.org
e24.ee folklore.ee delfi.ee www.ut.ee www.delfi.ee ut.ee Teeme ISE: harilikud hoidised virtuaalsetest viljadest veebiarhiivi riiulil Jaanus Kõuts, Eesti Rahvusraamatukogu 17.09.2013 Kogemused http://archive.org
PowerPoint Presentation
 Maamaksu infosüsteem (MAKIS) Maksustamishind Talumistasud Andres Juss Maa-ameti kinnisvara hindamise osakonna juhataja 13.11.2018 MAKIS eesmärk Kõik omavalitsused kasutavad veebipõhist maamaksu infosüsteemi
Maamaksu infosüsteem (MAKIS) Maksustamishind Talumistasud Andres Juss Maa-ameti kinnisvara hindamise osakonna juhataja 13.11.2018 MAKIS eesmärk Kõik omavalitsused kasutavad veebipõhist maamaksu infosüsteemi
1 Keskkonnamõju analüüs Koostajad: Koostamise aeg: metsaparandusspetsialist Madi Nõmm algus: bioloogilise mitmekesisuse spetsialist Toomas
 1 Keskkonnamõju analüüs Koostajad: Koostamise aeg: metsaparandusspetsialist Madi Nõmm algus: 17.04.2018 bioloogilise mitmekesisuse spetsialist Toomas Hirse lõpp: 24.07.2018 Tabel 1. Objekti üldandmed Harjumaa
1 Keskkonnamõju analüüs Koostajad: Koostamise aeg: metsaparandusspetsialist Madi Nõmm algus: 17.04.2018 bioloogilise mitmekesisuse spetsialist Toomas Hirse lõpp: 24.07.2018 Tabel 1. Objekti üldandmed Harjumaa
Väljaandja: Keskkonnaminister Akti liik: määrus Teksti liik: terviktekst Redaktsiooni jõustumise kp: Redaktsiooni kehtivuse lõpp:
 Väljaandja: Keskkonnaminister Akti liik: määrus Teksti liik: terviktekst Redaktsiooni jõustumise kp: 0.02.2009 Redaktsiooni kehtivuse lõpp: 3.0.206 Avaldamismärge: Kiirgustegevuses tekkinud radioaktiivsete
Väljaandja: Keskkonnaminister Akti liik: määrus Teksti liik: terviktekst Redaktsiooni jõustumise kp: 0.02.2009 Redaktsiooni kehtivuse lõpp: 3.0.206 Avaldamismärge: Kiirgustegevuses tekkinud radioaktiivsete
ITI Loogika arvutiteaduses
 Predikaatloogika Predikaatloogika on lauseloogika tugev laiendus. Predikaatloogikas saab nimetada asju ning rääkida nende omadustest. Väljendusvõimsuselt on predikaatloogika seega oluliselt peenekoelisem
Predikaatloogika Predikaatloogika on lauseloogika tugev laiendus. Predikaatloogikas saab nimetada asju ning rääkida nende omadustest. Väljendusvõimsuselt on predikaatloogika seega oluliselt peenekoelisem
Microsoft Word - PKT_hindamine_soomullad_2011_LYHI
 Soostunud ja soomuldade orgaanilise süsiniku sisaldus ja vastavalt sellele 1:1 mullakaardi võimalik korrigeerimine Töö teostajad: Põllumajandusuuringute Keskuse Mullaseire büroo, kontaktisik Priit Penu
Soostunud ja soomuldade orgaanilise süsiniku sisaldus ja vastavalt sellele 1:1 mullakaardi võimalik korrigeerimine Töö teostajad: Põllumajandusuuringute Keskuse Mullaseire büroo, kontaktisik Priit Penu
Euroopa Liidu Nõukogu Brüssel, 24. september 2015 (OR. en) 12353/15 ADD 2 ENV 586 ENT 199 MI 583 SAATEMÄRKUSED Saatja: Kättesaamise kuupäev: Saaja: Eu
 12353/15 ADD 2 ENV 586 ENT 199 MI 583 SAATEMÄRKUSED Saatja: Kättesaamise kuupäev: Saaja: Eu") Euroopa Liidu Nõukogu Brüssel, 24. september 2015 (OR. en) 12353/15 ADD 2 ENV 586 ENT 199 MI 583 SAATEMÄRKUSED Saatja: Kättesaamise kuupäev: Saaja: Euroopa Komisjon 23. september 2015 Nõukogu peasekretariaat
Euroopa Liidu Nõukogu Brüssel, 24. september 2015 (OR. en) 12353/15 ADD 2 ENV 586 ENT 199 MI 583 SAATEMÄRKUSED Saatja: Kättesaamise kuupäev: Saaja: Euroopa Komisjon 23. september 2015 Nõukogu peasekretariaat
Стальной кубик находится под действием сил, создающих плоское напряженное состояние (одно из трех главных напряжений равно нул
 Surutud varda abiisus (nõtke) Enamai varda otsad kinnitatakse ühe (Joon.1) näidatud neja viisi. Üejäänud kinnitusviiside puhu on kriitii jõudu võimaik määrata üdiatud Eueri vaemiga kp EImin, (1) kus -
Surutud varda abiisus (nõtke) Enamai varda otsad kinnitatakse ühe (Joon.1) näidatud neja viisi. Üejäänud kinnitusviiside puhu on kriitii jõudu võimaik määrata üdiatud Eueri vaemiga kp EImin, (1) kus -
ANOVA Ühefaktoriline dispersioonanalüüs Treeningu sagedus nädalas Kaal FAKTOR UURITAV TUNNUS Mitmemõõtmeline statistika Kairi Osula 2017/kevad
 ANOVA Ühefaktoriline dispersioonanalüüs Treeningu sagedus nädalas Kaal FAKTOR UURITAV TUNNUS Factorial ANOVA Mitmefaktoriline dispersioonanalüüs FAKTOR FAKTOR Treeningu sagedus nädalas Kalorite kogus Kaal
ANOVA Ühefaktoriline dispersioonanalüüs Treeningu sagedus nädalas Kaal FAKTOR UURITAV TUNNUS Factorial ANOVA Mitmefaktoriline dispersioonanalüüs FAKTOR FAKTOR Treeningu sagedus nädalas Kalorite kogus Kaal
INIMESEÕPETUSE AINEKAVA ABJA GÜMNAASIUMIS Klass: 10. klass (35. tundi) Kursus: Perekonnaõpetus Perekond Õpitulemused: Kursuse lõpus õpilane: 1) mõista
 Kursus: Perekonnaõpetus Perekond Õpitulemused: Kursuse lõpus õpilane: 1) mõista") INIMESEÕPETUSE AINEKAVA ABJA GÜMNAASIUMIS Klass: 10. klass (35. tundi) Kursus: Perekonnaõpetus Perekond : 1) mõistab, kuidas ühiskonnas toimuvad muutused avaldavad mõju perekonna ja peresuhetega seotud
INIMESEÕPETUSE AINEKAVA ABJA GÜMNAASIUMIS Klass: 10. klass (35. tundi) Kursus: Perekonnaõpetus Perekond : 1) mõistab, kuidas ühiskonnas toimuvad muutused avaldavad mõju perekonna ja peresuhetega seotud
Microsoft Word - vundamentide tugevdamine.doc
 10 Vundamentide tugevdamine. 1. Vundamentide tugevdamise põhjused 2. Tugevdamisega seotud uuringud 3. Tugevdusmeetodid 3.1 Vundamendi süvendamine 3.2 Talla laiendamine 3.3 Koormuse ülekanne vaiadele 3.4
10 Vundamentide tugevdamine. 1. Vundamentide tugevdamise põhjused 2. Tugevdamisega seotud uuringud 3. Tugevdusmeetodid 3.1 Vundamendi süvendamine 3.2 Talla laiendamine 3.3 Koormuse ülekanne vaiadele 3.4
Õppeprogramm „vesi-hoiame ja austame seda, mis meil on“
 ÕPPEPROGRAMM VESI-HOIAME JA AUSTAME SEDA, MIS MEIL ON PROGRAMMI LÄBIVIIJA AS TALLINNA VESI SPETSIALIST LIISI LIIVLAID; ESITUS JA FOTOD: ÕPPEALAJUHATAJA REELI SIMANSON 19.05.2016 ÕPPEPROGRAMMI RAHASTAS:
ÕPPEPROGRAMM VESI-HOIAME JA AUSTAME SEDA, MIS MEIL ON PROGRAMMI LÄBIVIIJA AS TALLINNA VESI SPETSIALIST LIISI LIIVLAID; ESITUS JA FOTOD: ÕPPEALAJUHATAJA REELI SIMANSON 19.05.2016 ÕPPEPROGRAMMI RAHASTAS: